共找到2條詞條名為性細胞的結果 展開
- 性細胞
- 胚細胞
性細胞
性細胞
遺傳學和細胞生物學名詞,也作生殖細胞或配子,是單倍體細胞,它由行有性生殖的生物在特定的器官通過減數分裂產生。兩性生殖細胞通過配子結合產生合子。您身體中的每一個細胞都含有來自您母親的卵子和您父親的精子的一套染色體。留意人類的染色體,您會發現它們呈X形,男性染色體則呈Y形。當身體製造性細胞(精子或卵子,視性別而定)時,身體必須減少一半的染色體,使之進入性細胞。為此,身體會在一次細胞分裂時從兩套染色體中隨機挑選,而後再次細胞分裂時將染色體數目減半。因此,您身體製造的每個精子或卵子都是獨一無二的——它包含了您父母基因的不同混合。正因如此,即使是同一家庭中同父同母的兄弟倆,也可能在長相和行為上截然不同——這完全取決於父母在製造性細胞時隨機抽選了哪些基因(染色體)。
性細胞(germ cell)是多細胞生物體內能繁殖後代的細胞的總稱,包括從原始性細胞直到最終已分化的性細胞。此術語由A·恩格勒和K·普蘭特爾於1897年提出以與體細胞相區別。體細胞最終都會死亡,只有性細胞有延存至下代的機會。物種主要依靠性細胞而延續和繁衍。長期的自然選擇使每一種生物的結構都為其性細胞的存活提供最好的條件。
性細胞(germ cell)又稱起配子(gamete),包括精子和卵子,均為單倍體細胞,即僅有23條染色缽,其中一條是性染色體。多細胞生物體內能繁殖後代的細胞的總稱。包括從原始性細胞直到最終已分化的性細胞。物種主要依靠性細胞而延續和繁衍。
在單細胞生物群體中已有性細胞分化的跡象。如團藻科的雜球藻有4個較小的細胞失去分裂能力,專司運動和代謝,稱為營養個體,其餘28個有分裂能力,稱為生殖個體;團藻則在大多數小型營養細胞間出現了少數大的性細胞。
在個體發生中,性細胞在發育早期就被決定了。在一些動物中,其決定因子可以追溯到上一代卵細胞的卵質。例如果蠅卵的後部有一特殊的細胞質區域稱極質,其中富含核糖核酸(RNA)的小顆粒叫極顆粒。經過受精、卵裂,含有極顆粒的細胞稱極細胞,就是果蠅的原始性細胞。如果將原在後部的極質注射到卵的前部,可使預定發育為體細胞的細胞發育為性細胞。在馬副蛔蟲卵中也可以看到類似的情況。有證據表明,兩棲類性細胞可能也是以同樣的方式決定的。但還不了解在哺乳動物胚胎中,是什麼因素決定某些細胞發育成性細胞的。只知道性細胞被決定后,需通過遷移到達生殖腺的部位並在那裡分化。
性細胞可以分成孢子和配子兩類:
是不需配合的性細胞,通常是無性的,可由減數分裂或有絲分裂產生,見之於原生動物中的孢子蟲綱和植物中。
是需經配合成合子後方能發育的性細胞,也稱性細胞,由減數分裂或有絲分裂產生。產生配子的細胞稱配子母細胞。這是未分化的原始性細胞,可在雄性或雌性生殖腺中分別分化為精子和卵。在低等植物中,配子母細胞在大多數情況下直接構成稱作配子囊的性器官。例如在原植體植物(Thallophyte)中的同形配子囊產生同形配子;異形配子囊產生異形配子等。
施旺和許耐登來說細胞仍然主要是一種結構成分,但在19世紀功年代其他學者已經強調了細胞的生理 功能,尤其是發育和營養功能。隨著對細胞及其組成成分(特別是細胞核)的了解日益深入,“細胞學說”這一概念的意義也逐漸發生了變化。許耐登的學說直接促進了區分動植物細胞的非常活躍的研究工作。1852年Remak(1815-1865)指陳,蛙卵是細胞,在發育中的蛙胚中新細胞是由原已存在的細胞分裂產生的。他著重駁斥了自由細胞形成學說。在這一點上他得到了微爾科(1855)的支持,後者指出,就許多正常的和病態的動物及人類組織來看,每個細胞都是從原已存在的細胞分裂而來。他斷言:“作為一般原則,任何種類的發育都不是從頭開始,因而(必須)否定(自然)發生學說,不論是在個別部分的發育史上還是在整體生物的發育史上”(Virchow,1858:54)。Kolliker以及其他一些植物學家也在同一時期得出了同樣的結論,然而,由於許耐登的權威性而延遲了這結論在植物學界被普遍接受。達爾文在1868年(II:370)還拿不準自由細胞形成究竟是否可能。後來微爾科的格言“細胞來自(原先存在的)細胞”,(Omnis cellula e cellula)雖然終於被每個人接受,然而那個時候對細胞(尤其是核)分裂過程的細節還並不清楚(見下文,“有絲分裂”)。
由於有了對細胞的這種新的解釋,重新考慮受精過程的時機已經成熟。如果軀體的一切部位都由細胞組成,則性腺(卵巢和睾丸)是否也是如此?雄性和雌性的“種子物質”究竟是什麼?它是否也由細胞構成?雌、雄性的性細胞有什麼區別?這一類早已構思過的問題當然並不是一開始就提了出來,然而這些問題必然是細胞學說的邏輯結果。後來越來越明顯只有等到闡明了受精過程中細胞的作用,真正有生命力的遺傳學說才能建立。正是在這幾十年中性細胞這個概念才脫穎而出。
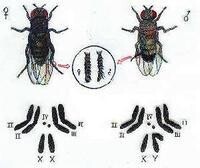
昆蟲的性細胞
多細胞生物體內能繁殖後代的細胞的總稱,包括從原始性細胞直到最終已分化的性細胞。此術語由 A.恩格勒和K.普蘭特爾於1897年提出以與體細胞相區別。體細胞最終都會死亡,只有性細胞有延存至下代的機會。物種主要依靠 性細胞而延續和繁衍。長期的自然選擇使每一種生物的結構都為其性細胞的存活提供最好的條件。
在單細胞生物群體中已有性細胞分化的跡象。如團藻科的雜球藻有4個較小的細胞失去分裂能力,專司運動和代謝,稱為營養個體,其餘28個有分裂能力,稱為生殖個體;團藻則在大多數小形營養細胞間出現了少數大的性細胞。
在個體發生中,性細胞在發育早期就被決定了。在一些動物中,其決定因子可以追溯到上一代卵細胞的卵質。例如果蠅卵的後部有一特殊的細胞質區域稱極質,其中富含RNA的小顆粒叫極顆粒。經過受精、卵裂,含有極顆粒的細胞稱極細胞,就是果蠅的原始性細胞。如果將原在後部的極質注射到卵的前部,可使預定發育為體細胞的細胞發育為性細胞。在馬副蛔蟲卵中也可以看到類似的情況。有證據表明,兩棲類性細胞可能也是以同樣的方式決定的。但還不了解在哺乳動物胚胎中,是什麼因素決定某些細胞發育成性細胞的。只知道性細胞被決定后,需通過遷移到達生殖腺的部位並在那裡分化(見生殖質)。
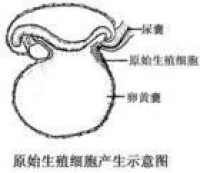
原始性細胞產生示意圖
精子中的半數含Y染色體(23,Y),半數含X染色體(23,X)。射出的精子雖有運動能力,卻無穿過卵子周圍放射冠和透明帶的能力。這是由於精子頭的外表有一層能阻止頂體酶釋放的糖蛋白。精子在子宮和輸卵管中運行過程中,該糖蛋白被女性生殖管道分泌物中的酶降解,從而獲得受精能力,此現象稱獲能(capacitation)。精子在女性生殖管道內的受精能力一般可維持1天。
從卵巢排出的卵子處於第二次成熟分裂的中期,並隨輸卵管傘的液流進入輸卵管,在受精時才完成第二次成熟分裂。若未受精,於排卵后12~24小時退化。
動、植物受精作用的實質是父本和母本的性細胞(配子)互相融合,這兩個配子在形成新的合子上各自作出了同樣貢獻,而且關鍵過程是兩個配子的細胞核相互融合,這些觀點到了1844年左右已經逐步確立並被有關學者普遍接受。人們的注意力便開始轉移到細胞核上。細胞核是不是就像後生論者所設想的,僅僅是一團無定形的胚樣物質、也許只是在融合時才激發了卵細胞的發育過程?或者是細胞核雖小,卻具有嚴密結構,這肉眼不可見的顯微結構是否就是受精作用之後的一切非常精確並具有特異性的發育過程的關鍵?如果把細胞核僅僅看作是細胞發育和細胞分裂的引發物,就會認為它在完成了這一任務后就會被溶解掉,在新的細胞分裂之前或至少是在配子形成之前再重新形成。由於19世紀後半期的細胞學家所接受的都是生理學家或胚胎學家的教育訓練,他們的側重點是發育問題,因而用不著關心細胞核的連續性。他們很少過向性狀是怎樣從親代傳遞到子代的遺傳學問題。
1875-1880年之間由於Balbiani,van Beneden,Flemming。Schleicher,Strasburger等五位學者在細胞分裂過程中能夠不斷追蹤其全部進程,因而“自由細胞形成”或細胞核“重新”形成這種信念的最後殘餘才得以完全肅清。這幾位學者論證了。
三項重要事實:
(1)在細胞分裂之前細胞核開始分裂;
(2)細胞核物質的變化具有正常順序(見下文);
(3)細胞核分裂和細胞分裂的基本現象在植物界和動物界都是相同的。
越來越明顯的是,細胞核的作用不單是生理性的(即純粹物理意義上的作為細胞分裂的引發物)。它是具有嚴密組織的、很可能是按特定格局構成的結構。這種格局的實質此後一直是細胞學者不斷關心的問題,仍然沒有求得最後答案。關於這方面研究進展的特點是分析越來越細。所涉及的步驟是從完整個體轉向細胞,從整個細胞轉向細胞核,現在則從完整的核轉向到它的主要結構成分、染色體。
有圓形外觀、偏心的核、以及充滿顆粒的細胞質,這些都是原始性細胞所具有的典型之形態特徵。魚類的第一個代理孕母。然而,為了獲得具有功能的配子,仍需努力把原始性細胞轉變成為精子和卵子的技術找出來。日本東京海洋大學的吉崎悟朗教授及其研究團隊曾以櫻鮭為代理孕母,在櫻鮭的腹膜腔內移植虹鱒的原始性細 還是多產的物種,只需一次受精,就可獲得數以千計,含有綠色螢光蛋白標記的原始性細胞的轉殖基因胚。一般而言,在魚類生殖隆起中的原始性細胞的數目很少,若要進行移植或體外培養實驗時,多產就成為一個很有利的條件,而虹鱒正好具有這個優點。流式細胞儀可以用來純化被綠色螢光蛋白標記的原始性細胞。從孵化期的胚上切下來的生殖隆起,經胰蛋白酉每溶解分散,透過流式細胞儀可分類成為被綠色螢光蛋白標記的細胞,和不被綠色螢光蛋白標記的細胞。前者直徑大約20微米,是一種具胞,藉由這個方法從櫻鮭生產虹鱒子代,這是世界上魚類的第一個代理孕母研究成功的報導。又為追蹤外源的原始性細胞在代理孕母體內的表現,研究人員先利用綠色螢光蛋白基因,把原始性細胞標記,然後再把孵化胚生殖隆起內的原始性細胞植入代理孕母。被植入到代理孕母腹膜腔內的原始性細胞,會朝向代理孕母的生殖隆起移動並形成群體聚落。到後來,來自供核者虹鱒的原始性細胞,在不同種(櫻鮭)的代理孕母的生殖腺中增殖,並分化為成熟的卵子和精子,再透過這些製造的配子受精,可生產出許多來自供核者的活仔魚。

基本信息
- 中文名
- 性細胞
- 外文名
- sex cell
- 別名
- 生殖細胞、配子
- 分裂方式
- 減數分裂或有絲分裂
- 種類
- 精子和卵細胞
- 特點
- 單倍體