染色體倍性
染色體倍性
染色體倍性(chromosome ploidy )是指細胞中包含的染色體組數或基因組數。如單倍體染色、多倍體染色。
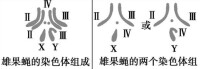
染色體倍性
由相同來源染色體組形成的多倍體稱為同源多倍體,由不同來源不同染色體組形成的多倍體稱為異源多倍體。此外,具有不成套染色體組的細胞或個體稱為非整倍體或異倍體(見染色體畸變)。
許多植物的染色體組包含著若干個祖先種(基本種)的染色體組。基本種的染色體組稱為基本染色體組,另以x表示,它所包含的染色體數稱為基數。相應地把包含若干祖先種的染色體組的物種稱為複合種,它所包含的染色體組數是2n=2∑x。複合種也常稱為多倍體物種。例如普通小麥通常稱為二倍體(2n)植物,同時又是異源六倍體植物,它的體細胞中的染色體數是2n=6x=42。
一般的二倍體生物在有性生殖過程中通過生殖母細胞的減數分裂使原來二倍體的染色體數目減半成為單倍體數。單倍體的雌、雄配子受精后形成二倍體的合子。在二倍體生物的生活史中,二倍體的體細胞與單倍體的性細胞形成正常的交替。
在動物界里生殖母細胞減數分裂后直接形成單倍體的卵子和精子。在植物界里二倍體孢子體的孢子母細胞經減數分裂所形成的是具有單倍染色體數的孢子,孢子通過有絲分裂形成雌、雄配子體,再由配子體產生卵子和(或)精子,精卵受精后產生二倍體的合子,合子再發育成孢子體。所以在植物中二倍體的孢子體與單倍體的配子體在個體生活史中形成明顯的世代交替。
真核生物在有性生殖過程中單倍體和二倍體的交替出現是生物界的普遍現象。多數低等的真核生物如黴菌的營養體是單倍體,少數種類如啤酒酵母的營養體是二倍體。在這些基本上是單細胞的生物中,營養體細胞和配子細胞之間往往沒有嚴格的區分,同時也容易得到它們的多倍體。在比菌藻植物更為高等的植物中使原來不能營獨立生活的世代變為能營獨立生活的世代便比較困難。比利時學者E.L.馬沙爾等在1911年首先用無性繁殖的方法使地錢(Mar-chantia polymorpha)的二倍體孢子體再生成為能營獨立生活的二倍體配子體。德國學者R.von韋特施泰因在1924年用同一方法得到葫蘆蘚(Funaria hygrometrica)的八倍體孢子體。這些是最早利用生物個體生活史中的染色體倍性交替現象來改變染色體組數的例子。
高等的顯花植物的孢子體已發展成為個體生活史中能營獨立生活的主體,而配子體反而成為隱蔽的、短時間的和寄生性的。印度學者S.古哈和S.C.馬赫什瓦里在1964年首先用花藥離體培養技術使曼陀羅的單倍體小孢子發育成植株,也就是使配子體轉變成孢子體,70年代以來用未授粉的子房培養也較易誘導產生單倍體,這就使人工大量製造單倍體的孢子體在高等植物中成為可能,在自然界中由孤雌(或孤雄)生殖所產生的單倍體則是極個別的。
在種子植物中胚乳是胚囊的兩個極核與一個精核通過雙受精形成的三倍體組織。通過組織培養方法,也能夠使胚乳細胞發育成為三倍體植物。
動物界中的膜翅目昆蟲蜜蜂的受精卵發育成二倍體的雌蜂和工蜂,而未受精的卵則發育成單倍體的雄蜂。在雄蜂的減數分裂的第一次分裂後期,各個染色體都集中在一極,只形成一個未減數的核,第二次分裂時才將子染色體均等地分向兩極,使所形成的兩個精子仍維持單倍體的染色體數。
正常的有絲分裂使生物的體細胞在增殖過程中維持恆定的染色體數。這是通過脫氧核糖核酸(DNA)的複製和隨之而進行的染色體、細胞核及細胞的依次分裂而實現的。只要其中某一步驟受到干擾,就會造成有絲分裂的各種不正常現象,從而導致染色體倍性的變化。如果只有DNA的複製,而沒有相應的染色體分裂,並且細胞核停留在有絲分裂的初期狀態,其結果就形成像果蠅等的唾腺細胞中的巨大多線染色體。
如果被抑制的不是染色體分裂而是細胞核分裂,那麼每一細胞中的染色體數目將成倍地增加,從而形成不同倍性的內多倍體,這種分裂方式稱為內有絲分裂。內多倍體現象在動物中(包括人類在內)是相當普遍的。人在11~20歲時肝細胞中常出現四倍體,到21歲以後就出現八倍體。家蚊(Culex pipiens)幼蟲的腸壁細胞核也存在內多倍體,它的二倍體染色體數是6,而腸壁細胞中的數目有12,24,48,96,甚至達到192。植物中在果實永久組織中也有8n,甚至多達32n的多倍性細胞。
如果有絲分裂只進行到核分裂而沒有細胞的分裂,結果就會形成多核細胞,如花藥絨氈層細胞等。一個細胞內的兩個核融合而使原來的二倍體細胞成為四倍體的現象在植物界是相當普遍的,也存在於許多動物中;一般認為這是多倍體細胞的主要來源。植物的愈傷組織中經常通過這種方式產生多倍體細胞。
如果這樣形成的多倍體細胞並不嚴重干擾器官分化,它們就會導致多倍體生物的產生。植物的細胞分化的程度較低,許多能夠分裂的細胞還保持了發育上的全能性,因此增加了由內多倍體細胞產生多倍體植物的可能性。高等的顯花植物中約有一半物種是多倍體(複合種)。雌雄同體的低等動物中也有多倍體生物。
荷蘭學者H.德·弗里斯在1886~1904年間所發現的巨型月見草後來經鑒定是同源四倍體。天然的同源多倍體物種不但在動物中極為罕見,即使在植物界中也不多見。
異源多倍體在植物界中相當普遍,丹麥學者O.溫厄在1917年提出有關異源多倍體的來源的學說,認為種間雜交后再經染色體加倍是植物物種形成的一個重要途徑。
種間雜交和雜種染色體的加倍可以在短時間內完成,所以有可能在自然界中觀察到這類新物種的出現和擴展,而且可以對它進行人工控制。C.L.赫斯金斯在1931年報道了大米草新物種(Spartina tonnsenolii)的出現和發展過程。大米草新種於1870年在英國南部海邊發現,到1902年擴大到幾千英畝,1906年出現在法國海岸,以後分佈到世界各地。1963年由中國植物生態學家仲崇信自英國將大米草引入中國,到70年代後期已成為南北海濱牧場的一個重要牧草。經研究認為這一新種是由具有56個染色體的歐洲本地大米草(S.stricta)與具有70個染色體的從美洲引入的美洲大米草(S.alterniflora)天然雜交而形成的異源多倍體,它的染色體數是126,是它的兩個親本物種的染色體數之和。
顯花植物中有許多物種是多倍體(見表)。在單子葉植物中,多倍體物種佔90%以上的有天南星科(Araceae)、鳶尾科(Iridaceae)、龍舌蘭科(Agavaceae)、蘭科(Orchidaceae)和莎草科(Cyperaceae)。禾本科中的箣竹族(Bambuseae)中幾乎全部物種都是多倍體。
首先發現藥物秋水仙素對有絲分裂產生影響的是義大利學者B.佩爾尼切,他在1889年描述了狗腸胃內壁細胞的分裂中期被秋水仙素所阻抑,紡錘體被破壞,染色體的行動癱瘓,不正常的有絲分裂大量停滯在分裂中期。這種由秋水仙素引起的不正常的分裂稱為秋水仙素有絲分裂(簡稱C-有絲分裂)。1937年美國植物遺傳學家A.F.布萊克斯利用秋水仙素加倍曼陀羅等植物的染色體數獲得成功。1962年中國遺傳學家嚴育瑞以C-有絲分裂為指標,從有機汞農藥中篩選出富民農,並用以加倍水稻、黑麥、小麥-黑麥雜種等植物的染色體數獲得成功。美國植物遺傳學家L.F.倫道夫在1932年用高溫短期處理玉米受精后的果穗,使幼胚的細胞成為四倍體,從而得到整株或部分是多倍體的玉米植株。
相反地,當染色體不再進行複製,而核分裂和細胞分裂照常進行時,通過同源染色體的配對和體細胞減數又可使內多倍體細胞回復到二倍體水平。
一般說來單倍體的花最小,二倍體、三倍體、四倍體依次增大。在花卉植物中,鬱金香(Tulipa)、唐菖蒲(Gladiolus)、觀賞鳶尾(Iris tro-jana)等的選種歷史一再證明選擇大形花會得到多倍體。
一般就植物的營養體來說,單倍體比二倍體生長差,三倍體和四倍體較好或接近於二倍體;就種子器官來說,四倍體比二倍體大。對有性生殖來說,凡倍性是單數的都將因減數分裂受到干擾而不育。這是單倍體分佈的隨機性造成大量沒有生活力的非整倍體的大、小孢子的結果。對不以種子為收穫對象的植物來說,三倍體育種具有重要意義。在這方面的最成功的例子是三倍體甜菜和三倍體無籽西瓜。
以四倍體甜菜作母本,與二倍體品種相間種植,結果從四倍體植株上收穫的種子約有75%是三倍體。1953年日本首先推廣三倍體甜菜。到60年代初,在西歐,三倍體甜菜幾乎完全代替了原來的二倍體品種。日本廣泛種植的一個茶樹抗寒品種和一個桑樹抗寒品種都是三倍體,最好的兩個除蟲菊品種也是天然的三倍體。美國約有四分之一的蘋果品種是三倍體。瑞典的林木育種家發現生長速度比普通白楊幾乎快一部的三倍體白楊。無籽的香蕉是天然的三倍體。日本植物遺傳學家木原均首先運用三倍體不育的特點,在1951年培育成功三倍體無籽西瓜。
在異源多倍體育種方面,蘇聯學者Т.Д.卡爾佩琴科在1927年首次取得具有9對染色體的蘿蔔(Raphanus sativus)與具有9對染色體的甘藍(Brassica oleracea)雜交的F1子代,並且在F2子代中得到染色體自發加倍而形成的完全可育的蘿蔔-甘藍異源四倍體。
1937年發現秋水仙素可以使染色體加倍以後,異源多倍體就能通過人工誘發使染色體數加倍而得到。這些多倍體常具有優良的生產性能。例如,使小麥和黑麥的雜種染色體數加倍而形成的八倍體小黑麥具有抗逆能力強、穗大、籽粒蛋白質含量高和生長勢強等優點。不過也有結實率低、種子不飽滿等嚴重缺點。因此,這類人工異源多倍體原始品系還須經過育種程序克服缺點,發展優點,然後才能在生產上發揮作用。
1972年美國遺傳學家P.S.卡爾森等採用原生質體融合方法將煙草屬的粉藍煙草(Nicotiana glauca)和郎氏煙草 (N.langsdorfii)兩個物種的體細胞融合在一起並誘導成植株。由體細胞融合所形成的這一雜種在染色體數目、形態和生理特性上完全與通過有性過程產生的異源多倍體相同。這一方法有可能被用來克服遠緣雜交中的困難,從而得到種間和屬間、甚至科間的雜種,為多倍體育種開闢新的途徑。
1、染色體變異:光學顯微鏡下可見染色體結構的變異或者染色體數目變異。
2、染色體結構的變異:指細胞內一個或幾個染色體發生片段的缺失(染色體的某一片段消失)、增添(染色體增加了某一片段)、顛倒(染色體的某一片段顛倒了180o)或易位(染色體的某一片段移接到另一條非同源染色體上)等改變。
3、染色體數目的變異:指細胞內染色體數目增添或缺失的改變。
4、染色體組:一般的,生殖細胞中形態、大小不相同的一組染色體,就叫做一個染色體組。細胞內形態相同的染色體有幾條就說明有幾個染色體組。
5、二倍體:凡是體細胞中含有兩個染色體組的個體,就叫~。如.人,果蠅,玉米.絕大部分的動物和高等植物都是二倍體。
6、多倍體:凡是體細胞中含有三個以上染色體組的個體,就叫~。如:馬鈴薯含四個染色體組叫四倍體,普通小麥含六個染色體組叫六倍體(普通小麥體細胞6n,42條染色體,一個染色體組3n,21條染色體。),
7、一倍體:凡是體細胞中含有一個染色體組的個體,就叫~。
8、單倍體:是指體細胞含有本物種配子染色體數目的個體。
9、花藥離體培養法:具有不同優點的品種雜交,取F1的花藥用組織培養的方法進行離體培養,形成單倍體植株,用秋水仙素使單倍體染色體加倍,選取符合要求的個體作種。
1、染色體變異包括染色體結構的變異(染色體上的基因的數目和排列順序發生改變),染色體數目變異。
2、多倍體育種:
a、成因:細胞有絲分裂過程中,在染色體已經複製后,由於外界條件的劇變,使細胞分裂停止,細胞內的染色體數目成倍增加。(當細胞有絲分裂進行到後期時破壞紡錘體,細胞就可以不經過末期而返回間期,從而使細胞內的染色體數目加倍。)
b、特點:營養物質的含量高;但發育延遲,結實率低。
c、人工誘導多倍體在育種上的應用:常用方法——用秋水仙素處理萌發的種子或幼苗;秋水仙素的作用——秋水仙素抑制紡錘體的形成;實例:三倍體無籽西瓜(用秋水仙素處理二倍體西瓜幼苗得到四倍體西瓜;用二倍體西瓜與四倍體西瓜雜交,得到三倍體的西瓜種子。三倍體西瓜聯會紊亂,不能產生正常的配子。)、八倍體小黑麥。
3、單倍體育種:形成原因:由生殖細胞不經過受精作用直接發育而成。例如,蜜蜂中的雄蜂是單倍體動物;玉米的花粉粒直接發育的植株是單倍體植物。特點:生長發育弱,高度不孕。單倍體在育種工作上的應用常用方法:花藥離體培養法。意義:大大縮短育種年齡。單倍體的優點是:大大縮短育種年限,速度快,單倍體植株染色體人工加倍后,即為純合二倍體,後代不再分離,很快成為穩定的新品種,所培育的種子為絕對純種。
4、一般有幾個染色體組就叫幾倍體。如果某個體由本物種的配子不經受精直接發育而成,則不管它有多少染色體組都叫“單倍體”。
5、生物育種的方法總結如下:
①誘變育種:用物理或化學的因素處理生物,誘導基因突變,提高突變頻率,從中選擇培育出優良品種。實例——青霉素高產菌株的培育。
②雜交育種:利用生物雜交產生的基因重組,使兩個親本的優良性狀結合在一起,培育出所需要的優良品種。實例——用高桿抗鏽病的小麥和矮桿不抗鏽病的小麥雜交,培育出矮桿抗鏽病的新類型。
③單倍體育種:利用花藥離體培養獲得單倍體,再經人工誘導使染色體數目加倍,迅速獲得純合體。單倍體育種可大大縮短育種年限。
④多倍體育種:用人工方法獲得多倍體植物,再利用其變異來選育新品種的方法。(通常使用秋水仙素來處理萌發的種子或幼苗,從而獲得多倍體植物。)實例——三倍體無籽西瓜和八倍體小黑麥的培育(6n普通小麥與2n黑麥雜交得4n後代,再經秋水仙素使染色體數目加倍至8n,這就是8倍體小黑麥)。
基本信息
- 中文名
- 染色體倍性
- 外文名
- chromosome ploidy
- 拼音
- ransetibeixing
- 定義
- 指細胞中包含的染色體組數或基因組數
- 出處
- 符號
- n
- 所屬學科
- 生物學
- 所屬領域
- 細胞學
- 分類
- 同源多倍體,異源多倍體,異倍體