佩氏亞齒菌是一種不可食用真菌,為革菌目班氏菌科亞齒菌屬的一員。它子實體的下表面懸掛著很多細齒狀的突起用於產生孢子。該菌主要分佈在北美,歐洲。佩氏亞齒菌是菌根菌,同多種針葉樹形成共生關係。佩氏亞齒菌的子實體通常有漏斗狀帶白邊的菌蓋,潮濕的幼子實體會流出鮮紅的汁液,其中所含色素有著與肝素類似的抗凝血活性。佩氏亞齒菌的幼子實體不尋常的外表為它贏得了許多生動的別名,例如奶油草莓菌,出血牙菌,紅汁牙菌和魔鬼牙菌。儘管佩氏亞齒菌在幼期很容易分辨,隨著它的成長,會逐漸變成不起眼的褐色。

佩氏亞齒菌
美國真菌學家霍華德·詹姆斯·班克(Howard James Banker)在1913年第一次對佩氏亞齒菌做出科學描述。義大利人皮爾·安德里亞·薩卡多1925將它歸到齒菌屬(Hydnum),而沃爾特·亨利·思奈爾(Walter Henry Snell)和愛思特·阿米利亞·狄克(Esther Amelia Dick)在1956年將其歸到麗齒菌屬(Calodon)。所以,佩氏齒菌和佩氏麗齒菌都是指佩氏亞齒菌。佩氏亞齒菌在分類上屬於亞齒菌屬的魔菌系(stirps Diabolum)。魔菌系真菌有一些共同特徵:菌肉上有明暗相間的同心圖案;嘗起來有很強的辛辣味;氣味略甜;孢子呈橢圓形;非澱粉質(不能被梅爾澤試劑染色);不能被甲基藍試劑染色;被細齒狀突起覆蓋;菌絲中有鎖狀連接。根據亞齒菌屬幾個菌種DNA分析的結果,佩氏亞齒菌與銹色亞齒菌(Hydnellum ferrugineum)已及一種學名為Hydnellum spongiosipes的亞齒菌親緣關係最近。佩氏亞齒菌的種名peckii是為了紀念真菌學家查爾斯·霍頓·佩克(Charles Horton Peck)。該菌俗稱奶油草莓菌,出血亞齒菌,紅汁牙菌,佩克的齒菌,出血牙菌和魔鬼牙菌。

覆蓋菌蓋的細齒是產生孢子的特別結構
與所有能長出蘑菇體的真菌類似,
子實體(擔子果)是佩氏亞齒菌的生殖結構。子實體是當一定的環境溫度,濕度和營養條件滿足時從菌絲體長出的。佩氏亞齒菌是一種有柄類齒菌(stipitate hydnoid fungus),也就是說,它有菌柄和菌柄末端的菌蓋,並且類似齒菌,在菌蓋下面有齒狀,而不是鰓狀或孔狀的子實層(hymenium)。多個子實體生長得很近的子實體常常看起來像是融合在一起。子實體能長到10.5厘米高。新鮮潮濕的子實體會流出鮮紅奪目的黏稠汁液。菌蓋外圍外凸,越往裡越平,有時中心會稍稍凹陷。它被稠密的突起覆蓋,摸起來像毛氈或天鵝絨。隨著菌株變老,這些突起會逐漸脫落,成熟的樣本最後只剩下光滑的菌蓋。菌蓋為圓形或不規則形狀,一般4到10厘米寬。對融合的菌株,寬度可達20厘米。菌蓋最初為白色,顏色逐漸變深,受傷的地方會留下深褐色到黑色的不規則斑塊。成熟的菌蓋表面堅硬,富含纖維,多鱗和鋸齒,菌蓋上層呈灰棕色,質地有點像木頭。裡層菌肉為淡粉紅至棕色。菌蓋上密布著細長的齒狀物,呈圓柱形和錐形,不到5毫米長,越往邊緣越短,每平方毫米通常有3到5個。它們最初為白色帶一點粉紅,逐漸變成灰棕色。菌柄粗短且常常扭曲變形,在穿透地表的地方呈球狀,可伸入土壤數厘米深。儘管菌柄總長可達5厘米,1-3厘米寬,但只有0.1到1厘米在地表以上。菌柄上部覆蓋著跟菌蓋下表面相同的齒狀物。而菌柄下部則多毛,常常纏繞著地表的碎屑。子實體的氣味被形容為“輕微的讓人不快”,或者按班克最初的描述,像山核桃仁。

新鮮的菌株在“流血”
沉積的孢子呈棕色。用
光學顯微鏡可以看到更精細的結構:大體呈球形,表面覆蓋疣狀節,長5.0-5.3微米,寬4.0-4.7微米。孢子為非澱粉質,也就是說在梅爾澤試劑的作用下不會吸收碘。佩氏亞齒菌的細胞體(菌絲)也有很多特徵。構成菌蓋的菌絲平滑透明,壁薄,約3-4微米。乾燥時會萎縮,但低濃度的
氫氧化鉀溶液(2%)可使其復甦。菌蓋中的菌絲複雜交錯,大體縱向延伸。它們被分為胞狀區間(septa)並且有鎖狀連接(clamp connections)。鎖狀連接是連接前一個細胞的短分支,用於提供細胞核分裂產物的通道。擔子體(basidia),即含有孢子的子實層細胞,呈梅花形,帶四個孢子,長35-40微米,寬4.7-6微米。

佩氏亞齒菌
亞齒菌Hydnellum diabolus(在某些文獻中種名為中性的diabolum)有非常相似的外形,以至於有些人認為它跟佩氏亞齒菌是同一種真菌。Hydnellum diabolus有佩氏亞齒菌所沒有的辛辣氣味。兩種真菌的差異清楚地體現在他們的成熟樣本中:Hydnellum diabolus有一個不規則的粗柄,而佩氏亞齒菌的柄是被一個海綿質地的層加粗。除此之外,老的佩氏亞齒菌有一個平滑的菌蓋,而Hydnellum diabolus被有絨毛。一個近親種Hydnellum pineticola也能從新鮮潮濕的子實體中流出粉紅色的液滴。它普遍生長在北美東北的針葉樹下。Hydnellum pineticola味道不好,但不辛辣。它的子實體常單身,而不會融合生長。另外,與佩氏亞齒菌不同,它沒有球狀柄。
佩氏亞齒菌是一種菌根真菌,會與某種樹的根(稱為宿主)形成互利的共生(mutualistic)關係,它能給宿主從土壤中提取出的礦物質和氨基酸,而得到宿主固化的碳。這種菌的地下菌絲能在很多種樹的小根周圍長出一層鞘組織。這種密切結合對宿主尤其有利,被稱為外生菌根(ectomycorrhizal)。這是因為真菌能產生把有機物礦物質化的酶,從而促進對宿主營養的輸送。佩氏亞齒菌的外生菌根結構已被詳細地研究過。它們的特徵是一個密絲組織幔層(plectenchymatous mantle)。這是一層由很多相互平行的菌絲緊密排列而成的組織,
菌絲幾乎不分叉或相互重疊。這些菌絲和附著的土壤礦物微粒都包在一種凝膠狀的基質中。這些外生菌根菌絲能變成厚壁孢子(chlamydospores)來幫助真菌忍受不利環境。佩氏亞齒菌的厚壁孢子有其不同於其他班氏菌的特殊結構:內壁厚而平滑,外壁有放射狀分佈的疣。整體上外生菌根最顯著的特點是它們黑色的,有“碳化的外形”老化外層組織。真菌的大多數地下組織都集中在地表附近,像一個由密集的外生菌根和菌絲體組成的菌絲墊。
菌絲體可以延伸到離子實體很遠的地方,有的可達337厘米。
分子遺傳學技術已被應用於保存包括佩氏亞齒菌在內的有柄類齒菌。雖然傳統上真菌分佈是根據子實體決定的,但這個方法有一個很大的缺點,就是子實體並不是每年都會長出來,而沒有子實體並不能說明土壤中不存在菌絲體。更現代化的技術通過聚合酶鏈反應確定土壤中是否有真菌DNA,從而緩解了探測真菌菌絲體存在與分佈的爭議。

佩氏亞齒菌可單身,散生或聚生
佩氏亞齒菌的子實體可以單生,散生或者聚生在針葉樹林地表,常常與
苔蘚和脫落的松針混在一起。佩氏亞齒菌是一種晚期(late-stage)真菌,在以
北美短葉松為主的
針葉林,通常會在樹冠閉合后才開始結合成熟的宿主。還發現它們對山區或亞高山生態系統有偏好。這種真菌在北美有廣泛分佈,特別是太平洋西北沿岸;北至阿拉斯加,東至
北卡羅來納。在美國華盛頓州的普吉特海灣地區,發現它與花旗松,杉和
鐵杉結合。在俄勒岡海岸地區的黑松下也搜集到了這種真菌。除了北美,它在歐洲也分佈較廣,有記錄的地區有義大利,德國和
蘇格蘭。在前面提到的這些地區,這種真菌比較普遍,但在其他某些歐洲國家則越來越罕見,例如挪威,荷蘭和捷克。這些地區逐漸惡化的污染狀況是導致真菌減少的可能原因之一。2008年在伊朗和2010年在韓國的發現是第一次在北美和歐洲之外發現該菌種。
雖然佩氏亞齒菌的子實體很像“點綴著
草莓果醬的丹麥糕點”,而且亞齒菌屬一般無毒,它們卻因為極其苦澀的味道而無法食用,這種苦味即使是在乾燥的樣品中也仍然存在。佩氏亞齒菌和其他亞齒菌的子實體是一種寶貴的染料。無媒染劑(mordant)時呈米色,用不同的媒染劑可以使它呈現不同色調的藍色和綠色。
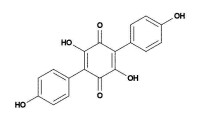
阿托門丁的分子結構式
化驗佩氏亞齒菌的提取物,可以發現存在一種有效的
抗凝血劑阿托門丁(atromentin,化學式:2,5-二羥基-3,6-二(4-羥基苯基)-1,4-
苯醌),它有著與著名的抗凝血劑肝素(heparin)相似的生物活性。阿托門丁還具有抗菌活性,它能抑制
肺炎鏈球菌中的烯醯-醯基載體蛋白還原酶(對脂肪酸合成必不可少)。最新研究表明,阿門托丁(atromentin,化學式:2,5-二羥基-3,6-二(4-羥基苯基)-1,4-苯醌)對聯三苯類成分vialinin A對TNF-α增殖有很強的抑制活性,具有良好的
抗病毒、抗腫瘤、抗菌、消炎、止血及子宮收縮等藥理活性,是一種天然的強抗氧化劑.
