稻大鼠是北美洲一種半水生的嚙齒類動物,它們主要分佈在美國東部及南部。它們一般出沒於潮濕的環境,如沼澤及鹽沼。它們重40-80克,屬於中等身型,外觀像黑鼠及褐鼠。稻大鼠是夜間活動的,以莎草及草來築巢,有時巢會有通道。它們的食性很多樣化,會吃植物、真菌及多種動物。倉鴞等都會掠食稻大鼠,在野外的稻大鼠一般都不能活過一年。多種的寄生蟲會寄生在稻大鼠之中,而它們也帶有一種可以感染人類的漢他病毒。它們並非保育的對象,但仍有一些群落受到威脅。
稻大鼠現正分類在稻鼠屬中。稻鼠屬分佈在美國東部至南美洲西北部,原先包含更多的物種,但都已被重置到其他地方。所有這些物種都被分類在稻鼠族中,稻鼠族是包含了很多個分支,超過100個物種的分類。稻鼠族以上的是棉鼠亞科及倉鼠科。除了庫氏稻鼠及棉鼠屬外,稻大鼠是美國唯一屬於棉鼠亞科。

稻大鼠
稻大鼠是由約翰·巴克曼牧師於1816年在南卡羅萊那州發現。他原先希望將稻大鼠分類在水䶄屬內,並將標本送往理察德·哈倫(Richard Harlan)及Charles Pickering處確認。連同另一個於新澤西州發現的標本,哈倫未獲Pickering的同意下將它們命名為小鼠屬的新物種,種小名 意為沼澤,是按其棲息而起的。於1854年,巴克曼牧師重新描述稻大鼠為,並認為它們最為接近
田鼠,且在喬治亞州及科羅拉多州發現其蹤跡。3年後,斯賓塞·富勒頓·貝爾德(Spencer Fullerton Baird)認為巴克曼牧師這樣分類錯誤,並建立了全新的稻鼠屬。稻鼠屬的命名是按它們吃稻米的習性。當時有認為稻鼠屬是一個完整的屬或為 的亞屬。但自1890年代起,一般已接納稻鼠屬為一個完全的屬,且與 有所分別。

稻大鼠側面圖
於1890年代,有幾個稻大鼠的亞種被描述:於1893年在科羅拉多州的、1894年在得克薩斯州的 及1898年在科羅拉多州以外地方的。柯林頓·哈特·梅里厄姆(Clinton Hart Merriam)於1901年確認 其實是另一物種,而 則差不多與指名亞種的 完全一樣。但於1918年,Edward Alphonso Goldman卻認為這些歸納到同一稻大鼠中。他分出了四個亞種,包括、 、 (包含)及。後來又加入了兩個亞種,包括松樹島(Pine Island)的 及桑尼伯島(Sanibel Island)的。於1955年,Claude W. Hibbard命名了稻鼠屬的新種,這是來自更新世地床的肯薩斯州,與現今稻大鼠只有牙齒上的少許分別。但是於1965年,Walter Dalquest根據在得克薩斯州發現,認為這個物種與稻大鼠分別不大,而將之降級成為亞種。梅里厄姆及Goldman發現一些中美洲物種,包括庫氏稻鼠,及一些不同形態都與稻大鼠有關。庫氏稻鼠分佈在得克薩斯州北部至最南端,與稻大鼠的分佈地鄰接。於1960年,Raymond Hall指在鄰接地方的一些標本其實是庫氏稻鼠與稻大鼠的中間形態,故此將庫氏稻鼠納入稻大鼠之內。不過後來卻發現庫氏稻鼠與稻大鼠實際上是有分別,後者較為細小,顏色較灰而非褐色,而且它們的染色體組型圖也有分別。自此以後,一般都將兩者看為獨特的物種,而於1994年更發現它們同時於得克薩斯州南部及墨西哥的塔毛利帕斯州生活。於1973年在佛羅里達礁島群發現了銀色稻鼠。究竟它們是獨立的物種或只是 的另一形態,仍然處於爭議階段。2005年利用微衛星DNA的研究發現它們與埃弗格拉德斯的稻鼠只有很少的基因分野,故認為它們只是不同的類群。這個類群可能是於約2000年前從大陸的稻鼠分支出來。於1989年就已描述的
亞種進行了一個形態定量分析,結果只分別出(包含了、 、 、及銀色稻鼠)及(包含了)。另一方面,有學者認為 及 為獨立的亞種,但卻將其餘的合併到,並將銀色稻鼠列為獨立的物種。

科羅拉多州的稻大鼠較其他地方呈紅色
稻大鼠是中等身型的
嚙齒類,外觀像黑鼠及褐鼠,但上身及下身的顏色有所不同。它們的毛厚而且短。上身一般呈灰至灰褐色,頭部的顏色稍淺,下身急轉為白色。
頰囊細小。耳朵與上身同色,耳前有一束淺色的毛。尾巴面呈深褐色,底呈淡黃色。護毛很長,尖端呈銀色。當它們游泳時,空氣會困在毛髮間,可以增加浮力及減少熱能流失。就像其他的稻鼠屬,雌鼠有8個額外乳房。稻大鼠的前腳有4趾,後腳有5趾,沒有絨毛。大部份腳上的墊都已退化,但在趾間有細小的蹼。佛羅里達礁島群的銀色稻鼠趾上有絨毛,但都已退化。這些都是水上生活的適應性。稻大鼠的毛色會稍按地理性出現變化:西方群落( )較東方( )的淺色,而科羅拉多州的一般較為紅色或黃褐色,科羅拉多州南部的( )則較中部的( )鮮色。佛羅里達礁島群的銀色稻鼠是呈銀色的,而另外兩個科羅拉多州形態 及,而是像 或褐色。於1989年,研究發現銀色稻鼠比 淺色,則較大陸群類深色,銀色稻鼠黃色毛的數量較少,但紅色毛則沒有分別。在群落間還有很多不同之處。
稻大鼠總長226-305毫米,尾巴長108-156毫米,後腳長28-37毫米,體重40-80克,雄鼠較雌鼠大隻。最大的個體於科羅拉多州及密西西比河三角洲東邊的美國墨西哥灣沿岸地區。稻大鼠的胃部像棉鼠類的沒有分開兩個部份,前部有腺上皮覆蓋。它們沒有膽囊,屬於稻鼠族的共有衍征。染色體組型圖包含了56個染色體,染色體短臂基數為60(2n=56;FN=60)。從性染色體可以分辨稻大鼠與庫氏稻鼠,但很難單以此來分辨其他稻鼠屬的物種。雖然稻大鼠沒有LINE-1反轉錄轉座子,但也會出現
X染色體去活化。在實驗室中也有出現融合或有額外
臼齒及毛色較淺的突變體。不正常臼齒似乎是由源自自體隱性的突變。稻大鼠的血細胞壓積約有50%,在嚙齒類中算是高的;這可能是稻大鼠在水下游泳是增加氧容量的一種適應性。
稻大鼠的頭顱骨
雄性稻大鼠的龜頭長而粗壯,平均長7.3毫米及闊4.6毫米,陰莖骨長6.6毫米。稻大鼠的陰莖構造複雜,陰莖骨兩側突出。陰莖外圍由細小的刺覆蓋,另有一道很闊及沒有刺的組織帶。陰莖上面的丘疹由小刺所覆蓋,是唯一與小嘯鼠屬及庫氏稻鼠相同的地方。位於陰莖末端凹陷位的尿道突有肉質的突;除了庫氏稻鼠及 外,其他的稻鼠類都沒有這個特徵。陰莖骨的深度比闊度長。中央的軟骨較側面的為大。雄性稻鼠類的附腺各有不同。雄性稻大鼠陰莖有一對包皮腺、兩對腹面前列腺、一對前面及背面前列腺。儲精囊末端部份不規則的褶疊。

稻大鼠的下頷骨
稻大鼠的頭顱骨很大及扁平,吻部較短及闊。鼻骨及前頜骨向後延伸越過淚骨、額骨及上頜骨的交接位。的吻部比科羅拉多州的扁平,而銀色稻鼠的鼻則較長。顴骨板很闊,前端有一結節。顴弧粗壯,有很多細小的顴骨。眶間最窄的部份向前傾。稻大鼠的顱腔很窄,由窄的頂間骨圍邊。及 的頭顱骨是最大及最闊的,西部的 的頭顱骨較為窄小。銀色稻鼠的頭顱骨也較為窄。
門齒孔闊達後頭的臼齒。齶很長,長至第三臼齒后。在近第三臼齒的位置有后側顎小凹。中翼窩上有延伸至前面的蝶齶窩。頭部的動脈高度進化。鱗狀骨沒有懸掛至鼓室蓋。乳突骨上有一些開孔。下頜骨的頦孔向兩邊開啟,而非如其他稻鼠類般向上開啟。上下咬肌嵴於第一稻齒下連接,並不會越過此位置。
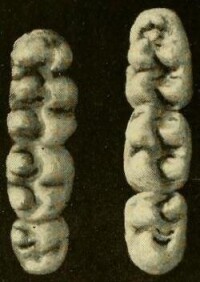
稻大鼠的上臼齒(左)及下臼齒(右)
稻大鼠的齒式是一隻上下門齒及三隻上下臼齒,一共16隻牙齒。上門齒髮育良好,咀嚼邊位於門齒垂直面后。臼齒有丘形齒冠及低冠齒型的。稻大鼠的上臼齒有兩行縱向的尖頂。第一及第二上臼齒呈卵狀。第一上臼齒的前尖頂並非由缺口來分開的,而是在中間有一虛位來分開唇面及舌面。唇面齒頂後有一冠。在第三臼齒,後面的尖頂已經退化及很難辨認。就像其他的稻鼠類,所有上臼齒都有一個內側齒根及兩個外側齒根。第一上臼齒一般會有多一個的外側齒根。第一下臼齒前端很圓,內側及外側小尖很難分辨。第二下臼齒很長,在兩個尖頂前有冠。臼齒前外邊有一明顯的脊。第三下臼齒與第二下臼齒差不多一樣長,同樣也在尖頂前有冠,但並不明顯,不過卻沒有前外邊的脊。第一下臼齒的前齒根很大,有1-2個細小后齒根。第二及第三下臼齒有兩個齒根,一般是外側及內側齒根,或是前端及後端齒根。
稻大鼠如其他稻鼠類般有12條肋骨。第一肋骨連接最後一節的頸椎及第一節的胸椎,是棉鼠亞科的共有衍征。第二及第三尾椎間有血管弧。它們沒有內上髁孔。
研究發現稻大鼠在惡劣環境下
腎上腺重量會增加至200%。它們在脫水下卻不能保存水份。它們如在被油污污染的水中游泳,死亡率亦會增加。殺死稻大鼠的輻射中位數是5.25戈瑞,氰化鉀的致死劑量是每公斤7.20毫克;兩項數值在倉鼠科內相對較低。有一項研究將稻大鼠放在受輻射污染的地區,它們卻沒有任何患病的徵象。實驗證實若稻大鼠較多曝露在日光下及有較多食物的供應,不論是成鼠或幼鼠,也會增強的性腺發育。若將雄鼠的
松果體移走或是餵食褪黑素,其睾丸會縮少退化。
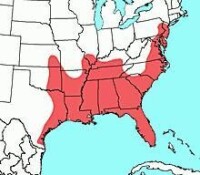
稻大鼠的分佈
稻大鼠現時分佈在美國東部及南部,東北至新澤西州南部,南至得克薩斯州東南部,東北至墨西哥塔毛利帕斯州。紀錄最北的曾在奧克拉荷馬州東部、肯薩斯州東南部、密蘇里州南部、
伊利諾伊州及肯塔基州南半出現,但卻絕跡於
阿巴拉契亞山脈。在科羅拉多州及喬治亞州的蘭喬拉布瑞亞時期地床曾發現稻大鼠化石,而已滅絕的 遺骸則是在得克薩斯州的威斯康辛冰期及Sangamonian地床和肯薩斯州的伊里諾冰期及Sangamonian地床發現。在佛羅里達礁島群,稻大鼠在大部份的下群島出沒,但卻絕跡於上群島。考古發現稻大鼠於全新世早期時,分佈得更北及更西,直達得克薩斯州中部、內布拉斯加州東部、艾奧瓦州西南部、伊利諾伊州中部、印地安納州南部、愛阿華州南部、西維吉尼亞州及賓夕法尼亞州西南部。大部份北方的考古位點都可以追溯至公元前1000年,並與玉米種植有關;但是在一些較為古老的洞穴內,稻大鼠則是與已滅絕的卡氏犰狳一同被發現,估計當時的氣候較為溫暖。可能於第四紀時的溫暖氣候,幫助稻大鼠向北擴展;而當氣候變得寒冷時,一些群落則因玉米種植而得而存活。稻大鼠在塔毛利帕斯州及得克薩斯州南部的分佈地與庫氏稻鼠鄰接,而在凱內迪縣、威拉西縣及卡梅倫縣,並塔毛利帕斯州東北部它們兩者並存。在實驗環境下,兩者並不能雜交,而基因分析亦沒有發現基因流或野生混種。相對於庫氏稻鼠,稻大鼠在群落內的遺傳變異較低,而在群落間的則較高,這可能是因它們的群落在近岸地區較成孤立所致。
稻大鼠棲息在多種環境,由海岸的鹽沼至山區的河流和林中空地。它們是半水生的,即大部份時間都會在水中生活,故很多時會在濕地出沒。它們喜歡長滿草及莎草的陸地,因為可以保護它們免受掠食者侵襲。它們也會在較干罕的高地出沒,因為這裡可以餵養幼鼠及在水漲時作為避難之用。稻大鼠可以橫渡海洋,如在維吉尼亞州德瑪瓦半島對出的海島的稻大鼠就可以橫渡島嶼間300米的海道。

倉鴞是稻大鼠的掠食者
稻大鼠在晚間活動,故此很難看見。它們會以莎草及草來築巢,巢約有13厘米大,會被築在垃圾下、近叢樹、洞穴內或水生植物之上。它們也會用沼澤鷦鷯、紅翅黑鸝、
麝鼠或圓尾麝鼠的巢。稻大鼠有時會製造很大通道或挖穴。它們是游泳的能手,在水下很易游多於10米,且當受到危險時,就會逃到水中躲避。在佛羅里達礁島群的稻大鼠有時會攀到樹上,但從未高於90厘米。稻大鼠非常整潔,經常擦洗全身,目的可能是要保持毛皮防水。它們對同類帶有攻擊性,打鬥時會發出高音叫聲。在密林中,它們的知覺感受範圍少於10米。當它們被釋放到其棲息地以外的地方,它們會向順風或逆風方向移動,來尋找適合的棲息地。多種動物都僧掠食稻大鼠。倉鴞是當中最為重要的;一項研究就發現倉鴞團粒中的脊椎動物遺骸就有97.5%是屬於稻大鼠的。其他的掠食者包括了鳥類(
白尾鷂及
橫斑林鴞)、蛇(
食魚蝮及其他)、
美國短吻鱷、食肉目(浣熊、
赤狐、
鼬屬及
臭鼬)。稻大鼠也有很多寄生蟲,體外的包括多種蜱蟎目、幾種虱毛目及兩種跳蚤,體內的包括多種
線蟲動物門及復殖目、一種舌形動物門及幾種球蟲亞綱。牙周病影響著稻大鼠的顎部;而它們也被用作研究牙周病的對象。維他命E、氟化物及碘化物可以保護患病稻大鼠的骨頭,高蔗糖的食性會增加稻大鼠患上牙周病的嚴重性。
稻大鼠的群落密度不會多於每公頃10隻。天氣會影響群落的動態;在佛羅里達大沼澤地,當泛濫將群落聚集起來時,密度可以多過每公頃200隻;在佛羅里達礁島群,密度則可以少於每公頃1隻。在路易西安那州的Breton島,雄鼠的活動範圍約為0.37公頃,雌鼠的則為0.23公頃。在科羅拉多州的研究發現,雄鼠的活動範圍平均為0.25公頃,雌鼠的則為0.33公頃。群落大小一般在夏天是最大的,而冬天則會減少,但在得克薩斯州及路易西安那州的群落大小則比較穩定。它們一般在冬天會變瘦。在得克薩斯州南部的群落大小每年的變化很大。在密西西比州海岸,研究發現風暴並不會令它們的群落減少;在得克薩斯州,泛濫亦不怎麼影響它們的密度。不過,另一項研究卻發現泛濫會大大影響
密西西比州的稻大鼠數量。
在稻大鼠北部的分佈地,它們一般會與草原田鼠一同出現,但卻沒有證據顯示它們互相爭競。在南部,剛毛棉鼠也會與稻大鼠一同出沒,水平線會影響著它們的相對數量。棉鼠主要是日間活動的,與稻大鼠的生態位有所分野。

互花米草是稻大鼠的食物
稻大鼠會以植物及動物為食物,主要的食物會隨季節而變更。植物食物包括
大米草、鹽角草屬、磨擦草屬及披鹼草屬等;它們主要吃種子及肉質部份。研究發現它們喜歡吃
互花米草的莖內組織,可能是該部份含有較少二甲基巰基丙酸內鹽。稻大鼠是稻米的主要害蟲,會吃剛種植的稻米。它們有時也會吃內生菌屬。動物食物包括昆蟲、
招潮蟹及蝸牛,不過也會吃其他動物,如魚類、蛤、幼生的
地圖龜及
錦龜。它們會吃麝鼠、白足鼠屬及麻雀的腐肉。稻大鼠也會吃沼澤鷦鷯及海灘雀的雛鳥及鳥蛋,特別對海灘雀具有攻擊性,令它們不敢在
燈心草屬上築巢。在
北卡羅來納州的島嶼,稻大鼠會吃加拿大燕鷗的鳥蛋。也有見過它們在喬治亞州吃
短吻鱷的蛋。研究發現稻大鼠會同化88%-95%的食物。若只吃大米草、招潮蟹或向日葵的種子,它們的體重就會減輕。它們並沒有儲藏行為。在高地的稻大鼠主要吃水生植物及動物,也會吃一些高地植物。
稻大鼠主要於夏天繁殖。有些指它們會於冬天完全終止繁殖,但卻在維吉尼亞州的仍會在冬天繁殖。在得克薩斯州及維吉尼亞州,雌鼠的繁殖活動較雄鼠的少。在南部分佈地,它們在夏天最溫暖的日子繁殖較少。動情周期歷時6-9日,平均則為7.72日。在生下一胎后也會出現動情。稻大鼠的交配行為與褐鼠的相似。在交配前,雄鼠會跟在雌鼠后。雄鼠接著會重複爬上及跳下雌鼠,並非每次爬上都會交配。侵入只會歷時0.25秒,但在交配時侵入的時間會較長。縱然雄鼠已經完成交配,只要有新的雌鼠出現,它們也可以立即再進行交配。部份原因是雌鼠帶有抗拒性,雄鼠射精的頻率較低。
稻大鼠的妊娠期為25日,每胎可以有1-7隻幼鼠出生,平均有3-5隻。雌鼠一年可以生6胎。初生幼鼠重3-4克,不能看見及沒有毛。出生的雄鼠與雌鼠比例相約。後期
外耳會打開,第一天就可以見到它們的爪。第二天它們就會抓東西,到了第三至第五日,就會長出須及眼廉。期后的兩天,乳頭及門齒就會長出,幼鼠此時會變得較為活躍。介乎八至十一日大,幼鼠會打開眼睛,長出毛皮,並會開始吃固體食物。第十一至十二日大就會斷奶。不同年齡幼鼠的體重也會按地理性而有所差異。它們要到60日大來達至性成熟。野生的稻大鼠一般只可以活少於1年;平均壽命則為7個月。
稻大鼠一般對人類並不重要,故此對它們的研究較其他北美洲嚙齒目為少。它們曾被建議作為研究其他嚙齒目感染的模式生物。稻大鼠在美國是牛軛湖病毒的主要帶菌者,也是漢他病毒的第二普遍的感染者。約有16%的稻大鼠受到感染,尤其是較年長及重的雄鼠。病毒可以經由稻大鼠傳播。在它們的唾液及尿液中也含有病毒,人類接觸這些排泄物后就有可能感染。兩種相關的漢他病毒分吃在宏都拉斯及墨西哥西部的庫氏稻鼠中發現。在林鼠屬中發現的沙狀病毒科也可以在稻大鼠中發現。
2009年IUCN紅色名錄將稻大鼠列為無危,原因是它們普遍、廣泛分佈及數量穩定。在佛羅里達礁島群的稻大鼠較為稀有,數量更在下降,原因受到與黑鼠競爭、被家貓掠食、失去棲息地及失去遺傳變異的威脅,現正被認為是瀕危。
