漢坦病毒
有包膜分節段的負鏈RNA病毒
漢坦病毒歸屬布尼亞病毒科,是一種有包膜分節段的負鏈RNA病毒,基因組包括L、M、S 3個片段,分別編碼L聚合酶蛋白、G1和G2糖蛋白、核蛋白。
漢坦病毒腎綜合征出血熱(HFRS)由漢坦病毒引起的一種自然疫源性疾病,是嚴重危害我國人民健康的病毒性疾病之一,為《中華人民共和國傳染病防治法》規定的乙類傳染病。
漢坦病毒
漢坦病毒包括引起腎綜合征出血熱(HFRS)的漢灘病毒(Hantaan virus,HTNV)、漢城病毒(Seoul virus,SEOV)、普馬拉病毒(Puumala virus,PUUV)、多不拉伐病毒(Dobrava virus,DOBV),引起漢坦病毒肺綜合征(HPS)的無名病毒(Sin Nombre virus,SNV)、紐約病毒(New York virus,NYV)、污黑小河溝病毒(Black Creek Canal virus,BCCNV)、牛軛湖病毒(Bayou virus,BAYV)、安第斯病毒(Andes virus,ANV)以及與人類疾病關係尚不清楚的一組病毒,如希望山病毒(Prospect Hill virus,PHV)、泰國病毒(Thailand virus,THAIV)、圖拉病毒(Tula virus,TULV)、索托帕拉雅病毒(Thottapalayam virus,TPMV)、哈巴羅夫斯基病毒(Khabarovsk virus,KBRV)、El Moro Canyon病毒(ELMCV)、Rio Segundo病毒(RIOSV)、島景病毒(Isla vista virus,ISLAV)、Muleshoe病毒(MULEV)、Bloodland lake病毒(BLLLV)、Rio Mamore病毒(RMV)、Topografov病毒(TOPV)等。
漢坦病毒實驗診斷方面的研究,主要集中於重組抗原的應用和實驗診斷方法的快速、敏感和特異。 F.Elgh等採用PUU病毒重組核蛋白作為抗原,與乳膠連接進行乳膠微粒凝集試驗,用於漢坦病毒病的快速血清學診斷,與採用PUU病毒重組核蛋白作為抗原的ELISA相比,特異性為90%,敏感性為94%。Jiro Arikawa等將桿狀病毒表達的HTN、SEO、PUU病毒核蛋白用於ELISA,至少應用2種重組抗原(HTN和PUU或SEO和PUU),可以用於漢坦病毒感染的血清學監測。桿狀病毒表達N端缺失的HTN或SEO病毒核蛋白作為免疫熒光試驗(IFA)的抗原,可以區分HTN與SEO病毒感染。重組核蛋白和N端缺失的核蛋白用於ELISA和IFA,提供了針對漢坦病毒感染的快速、敏感、安全的診斷方法。H.Kallio-Kokko等將桿狀病毒表達的PUU病毒核蛋白用於IgG和IgM檢測,將大腸桿菌表達的PUU病毒核蛋白用於IgM檢測,敏感性達100%,部分表達的核蛋白用於IgG檢測,敏感性較低(70%)。他們還報道了在PUU病毒感染的急性病例中,2/3的病例可以採用RT-PCR試驗,從病人的血或尿中檢出病毒RNA。T.Tomiyama等將高密度正性顆粒包被純化的漢坦病毒抗原,採用高密度顆粒凝集試驗(HDPA)對病毒感染進行快速血清學診斷,對HTN病毒感染的檢測有較高的敏感性和特異性,對PUU、SN病毒的感染也有低水平的交叉反應。HDPA與IFA比較,敏感性相近,但比IFA 更為簡便快速。W.Irwin等報告了現場調查中免疫印跡試驗在鼠類病毒抗體檢測中的應用。邱建明等採用5’端生物素標記漢灘病毒特異性寡核苷酸探針,結合磁性分離技術及改進的異硫氰酸胍-酚一步法兩種方法提取病毒RNA,進行反轉錄套式PCR,用於檢測臨床HFRS病人血清。在7d以內病人血清的陽性檢出率為100%,8~14d病人血清的陽性檢出率為57.14%,15d后病人血清仍能檢測到22.73%陽性。擴增產物經打點雜交檢測證實為特異性擴增,這為早期確診HFRS病人提供了特異、敏感、快速、直接的診斷方法。
各國學者採用病毒與宿主的基因分析方法,研究漢坦病毒分離株之間或宿主動物之間親緣關係的遠近,以及病毒與宿主動物的共演化。
漢坦病毒基因分析
漢坦病毒
宿主動物基因分析
多採用細胞色素B基因分析方法。W.C.Black IV等採用微衛星DNA分析確定鹿鼠之間的基因關係,並研究了鼠類窩內漢坦病毒傳播的方式。
共演化過程
漢坦病毒
漢坦病毒

實驗室診斷
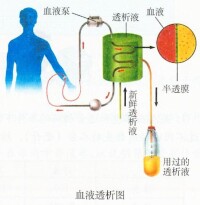
血液透析
HFRS是嚴重危害我國人民健康的病毒性疾病之一。自80年代初成功分離病毒以來,HFRS和漢坦病毒的研究取得了大量成果,尤其近年來滅活疫苗的研製成功,為有效預防本病創造了條件。但我們在病原學、實驗診斷、免疫病理和分子生物學等方面與國外研究尚有差距,仍存在許多有待解決的問題。隨著今後研究的深入,對本病認識的逐步加深,才能最終有效控制本病在我國的流行。
智利1995年發現首例HPS病人,1995年10月至1997年7月發病僅8例,1997年10~12月發病20例,涉及全國11個地區,平均發病年齡29.7歲,病死率61%。病毒基因分析表明,發病主要由Andes病毒引起。作者認為,HPS病例的增加與當地漢坦病毒宿主動物數量的增加有關。俄羅斯1978~1996年共發生HFRS91 639例,分佈於89個行政區中的61個,其中96.4%來自俄羅斯歐洲部分,3.6%來自亞洲部分,年平均死亡率分別為4.0/10萬和0.6/10萬。血清學和基因分型表明,俄羅斯至少存在6種血清型的漢坦病毒:HTN、PUU、SEO、TUL、KRB、TOP。比利時1995年10月至1996年12月發生HFRS 199例,季節分佈表現為冬春季的小高峰和夏秋季的大高峰,病人主要為PUU血清型感染。加拿大截止1997年11月共發生HPS 21例,分佈於3個西部省份,病死率33%,流行病學調查表明全國都有攜帶病毒的宿主動物分佈,而HPS發病與接觸鼠類的機會有關。日本從17個海港中的16個和3個飛機場中的2個檢出宿主動物攜帶漢坦病毒,作者提出應建立監測體系並採取相應預防措施,檢查人群病毒感染率。
氣候、鼠類棲息地條件、鼠類繁殖強度、種群構成等因素的變化影響鼠密度,而宿主動物病毒感染率隨著時間和地點的不同不斷發生變化。不同的鼠密度、宿主動物病毒感染率和與人群接觸的機會,影響疾病的暴發或散發。T.S.Chiueh等對台灣進行的血清流行病學研究表明,當地雖然沒有確診的HFRS病人,在宿主動物中卻存在漢坦病毒的感染,在與鼠類接觸的職業人群和慢性腎衰及發熱病人中,血清抗體陽性率較高。C.Ahlm等報告瑞典北部野生駝鹿具有較低的漢坦病毒感染率。Yun-Tai Lee等發現,韓國蝙蝠、棕頭鴉雀攜帶PUU病毒。O.A.Alexeyev等的研究表明,在PUU抗體陽性歐洲棕背中,大部分(74%)不能檢出病毒RNA或抗原,也不具有感染性。此外,為表示漢坦病毒株對Vero-E6細胞適應能力與宿主動物種類的關係,L.Ivanov等用鼠肺標本漢坦病毒抗原滴度、病毒分離成功率、病毒分離天數等計算適應指數,黑線姬鼠為0.32,大林姬鼠為0.17,東方田鼠為0.10,棕背為0.06。
主要臨床表現為,在4日左右的發熱、頭痛等前驅期癥狀后,出現以非心源性肺水腫和高病死率(52.4%~78.0%)為特徵的急性呼吸衰竭,重症3~7日死亡,生存者則很快恢復,無後遺症。其發病機制主要是病毒的直接致病作用,腎臟是早期原發性損傷器官,病毒是腎損傷的直接因素。
漢坦病毒實驗診斷方面的研究,主要集中於重組抗原的應用和實驗診斷方法的快速、敏感和特異。
多項報告對HFRS和HPS的後遺症進行了調查,研究表明兩種疾病的恢復病人與健康人比較,分別存在腎臟或肺功能的異常。M.Howard等對患有HPS的孕婦進行了調查,結果表明患有HPS的孕婦預后與其他HPS患者相同,患有HPS孕婦的胎兒與其他患有成人呼吸窘迫綜合征的孕婦的胎兒差異不大,研究中沒有發現SN病毒在人類垂直傳播的現象。
HFRS的發病主要是病毒直接作用所致,病毒血症及外周血單核細胞內病毒存在的時間一般 為7~10日或長,早期抗病毒治療可阻斷病理損傷、減輕病情、降低病死率。應用干擾素、利巴韋林,有肯定療效。7病日以內均可應用,療程5~7日。
應強調預防性治療,不同病期有不同的液體療法原則。以平衡鹽液為主,根據化驗檢查結果,適當調整其成分及用量。應積極糾正低蛋白血症。
美國國家公園突發漢坦病毒
美國加利福尼亞州度假勝地優勝美地國家公園宿營區近日出現致命漢坦病毒感染病症,截至9月7日致死人數已經上升至3人。
據悉,第3名死者來自美國西弗吉尼亞州,目前確認感染漢坦病毒的總人數已經達到8人,並且有可能繼續上升。
此前的2名死者分別來自美國加利福尼亞州和賓西法尼亞州。其他感染者均來自加利福尼亞州,公園工作人員稱他們的病情已經有所緩解。
據悉,美國衛生官員已向39個國家發出警告,稱今年夏天曾在優勝美地國家公園宿營的該國公民可能感染致命的漢坦病毒。
專家稱,大約有1萬人因6月到8月期間去過優勝美地國家公園而存在感染漢坦病毒肺綜合征的風險,其中約2500人來自美國以外的國家。目前尚無治癒該疾病的方法,但通過血液檢查及早發現可以極大地提高生存機率。
當地時間2021年6月7日,美國密歇根州衛生官員報告了首例人類確診感染漢坦病毒的病例。
雲南省發現漢坦病毒
2020年3月23日,雲南省臨滄市滄源縣一名外出務工人員途經陝西省寧陝縣時出現身體不適,於當日07:10搶救無效死亡。經採樣檢測,其漢坦病毒核酸檢測為陽性。
2020年3月24日,記者採訪了雲南省地方病防治所黨委書記左仕富。他表示,該病為乙類傳染病,近5年雲南已報告1231例漢坦病毒患者,其中1人死亡。