共找到5條詞條名為進化論的結果 展開
- 生物學術語
- 三聯書店傳的同名書籍
- 劉辰希演唱歌曲
- 電影
- 林俊傑作品
進化論
生物學術語
進化論是一種科學理論,指以史為鑒找出生命生存規律和發展方向,是對物種起源和發展的一種科學證明。21世紀,以分子鐘為基礎,以蛋白質PAM矩陣和BLOSUM的氨基酸矩陣為證據,在分子系統發生學和進化動力學的基礎上,繼續發展達爾文的進化論。
進化論的研究內容之一是進化生物學,研究合胞滋養層。
查爾斯·達爾文(1809-1882),英國生物學家、博物學家,進化論的奠基人,早期以地質學研究而聞名,而後又在動植物和地質方面進行了大量的觀察和採集,提出科學證據,證明所有生物物種是由少數共同祖先,經過長時間的自然選擇過程后演化而成。
宇宙中最原始的存在,並不是具有精神的事物、靈或神,而是具有生能的物質。這生能以進化方式,演進成生元,即細胞;這細胞便是萬物中一切生命的開始。
在西方思想史上,力持進化論觀點的是英國生物學家達爾文(1809-1882)。經過多年的探索,達爾文逐漸形成了一個系統的進化思想:生物界本來就存在著個體差異,在生存競爭的壓力下,適者生存,不適者被淘汰;物種所保留的有利性狀在世代傳遞過程中逐漸變異,經過性狀分異和中間類型消失便形成新物種。達爾文主張,生物界物種的進化及變異,系以天擇的進化為其基本假設:此外,並以性別選擇和生稟特質的遺傳思想來作輔助。1859年,達爾文的《物種起源》的出版,震動了整個學術界和宗教界,強烈地衝擊了《聖經》的創世論。達爾文的《物種起源》提出生物進化論學說,對宗教“神造論”和林奈與居維葉的“物種不變論”發起一場革命,震動當世。迄今,越來越多生物物化石發現,日新月異的基因遺傳學研究成果,已越來越有力證明了進化論的科學性。由於進化論違反《聖經》里的創世論,所以自問世以來,一直是宗教爭論的焦點,甚至在今天,篤信宗教者仍然宣稱進化論不可信。
早期理論
列子(約前450年—約前375年)為中國古代(戰國時期)著名的思想家,他在《說符》中說,“天地萬物,與我並生類也。類無貴賤,徒以大小智力而相制,迭相食,非相為而生之。人取可食者而食之,豈天本為人生之?且蚊蚋囋膚,虎狼食肉,非天本為蚊蚋生人、虎狼生肉者哉”。有人解釋為物種間的關係也絕不是所謂上天(神)目的性設計的結果,包括人在內的生物界只不過是自然發展的產物,人也非比其它動物更為高貴生靈。但此解釋也有爭議,因為列子學說屬於中國道家思想範疇,而道家和後來的中國道教並不完全否定“神創說”。
古希臘阿那克西曼德(約公元前6世紀)認為生命最初由海中軟泥產生。原始的水生生物經過蛻變(類似昆蟲幼蟲的蛻皮)而變為陸地生物。他推測,人最初是由另外一種動物產生的,其理由是別的動物都很快就給自己尋找食物,而只有人卻需要很長一段哺乳時期,如果人在起初就是像現代人這種樣子,那他是不能存留下來的。他認為人就是從魚產生的,這可能是古人從觀察人的胚胎和幼魚有某種相似而得出的結論。
古希臘亞里士多德(公元前384—前322年)在《動物志》中說,“自然界由無生物進展到動物是一個積微漸進的過程,因而由於其連續性,我們難以覺察這些事物間的界限及中間物隸屬於哪一邊。在無生物之後首先是植物類……從這類事物變為動物的過程是連續的……”,在《論植物》中說,“這個世界是一個完整而連續的整體,它一刻也不停頓地創造出動物、植物和一切其他的種類”。他認為生命的演化應該是這樣的途徑:非生命→植物→動物(這被後人稱之為“偉大的存在之鏈”),這也大致符合基於現代科學的認知。達爾文進化論中的微小變異的連續累積似乎也與亞里士多德的積微漸進的生命演化觀如出一轍。
宗教學特創論
中世紀的西方,基督教聖經把世界萬物描寫成上帝的特殊創造物。這就是神創論(創造論)。神創論表示整個自然界被創造出來是為了彰顯造物主的榮耀。
智慧設計論
智慧設計論又稱作目的論,它是一種神學論,由於智慧設計論一般來說是沒有神的。智慧設計論是說世界必然是由一個超自然的東西來創造並設計了(這些實體和)某些規則,造成了這些現象。這些現象的特徵主要可歸納為不可化約的複雜性、特定複雜性,以及宇宙萬物有序、符合規律。在細胞膜是脂融性弱酸被證明後幾經被否定,在基因生物學建立的PAM矩陣面前徹底被否定。
不變論
從15世紀後半葉的文藝復興到18世紀,是近代自然科學形成和發展的時期。這個時期在科學界占統治地位的觀點是不變論。當時這種觀點被牛頓和C.v林奈表達為科學的規律:地球由於所謂第一推動力而運轉起來,以後就永遠不變地運動下去,生物物種本來是這樣,以後也是這樣,其也被否認了。
活力論
到了18世紀下半葉,I.康德的天體論首先在不變論自然觀上打開了第一個缺口;隨後,轉變論的自然觀就在自然科學各領域中逐漸形成。這個時期的一些生物學家,往往在兩種思想觀點中入門徬徨。
例如林奈晚年在其《自然系統》一書中刪去了物種不變的詞句;法國生物學家G.-L.de布豐雖然把轉變論帶進了生物學,他一生都在轉變論和不變論之間徘徊。J.-B.de拉馬克在1809年出版的《動物哲學》一書中詳細闡述了他的生物轉變論觀點,並且始終沒有動搖。
18世紀末—19世紀後期,大多數動植物學家都沒有認真地研究生物進化,而且偏離了古希臘唯物主義傳統,墜入唯心主義。
“活力論”雖然承認生物種可以轉變,但是把進化原因歸於非物質的內在力量,認為是生物的“內部的力量”即活力驅動著生物的進化,使之越來越複雜完善。活力論缺乏實際的證據,只是一種唯心的臆測。
拉馬克主義
拉馬克主義,又稱用進廢退論,在活力論的影響下,最有名的活力論者就是法國生物學家拉馬克。19世紀前期出現的終極目的論或直生論,認為生物進化有一個既定的路線和方向而不論外界環境如何變化。後人把拉馬克對生物進化的看法稱為拉馬克學說或拉馬克主義,其主要觀點是:
(1)物種是可變的,物種是由變異的個體組成的群體。
(2)在自然界的生物中存在著由簡單到複雜的一系列等級(階梯),生物本身存在著一種內在的“意志力量”驅動著生物由低的等級向較高的等級發展變化。
(3)生物對環境有巨大的適應能力;環境的變化會引起生物的變化,生物會由此改進其適應;環境的多樣化是生物多化的根本原因。
(4)環境的改變會引起動物習性的改變,習性的改變會使某些器官經常使用而得到發展,另一些器官不使用而退化;在環境影響下所發生的定向變異,即後天獲得的性狀,能夠遺傳。如果環境朝一定的方向改變,由於器官的用進廢退和獲得性遺傳,微小的變異逐漸積累,終於使生物發生了進化。
拉馬克學說中的內在意志帶有唯心論色彩;後天獲得性則多屬於表型變異,現代遺傳學已證明它是不能遺傳的——拉馬克“用進廢退進化論”取材於現實觀察,推論於進化遺傳,它是“勞動創造人本身”的先聲。現代遺傳學所取材的觀察實驗既沒有確定宏觀進化的發生也不能確定後天獲得性狀不能遺傳(說明詞:“可能不會發生”)。進化論和遺傳論兩者的核心差異是:進化論認為,對生命來說,客觀的環境外因和主觀的生命內因的選擇是互相影響的;遺傳論則否定生命存在內因選擇,是為避開進化論洪流而構建的另一種命由天定論。
1858年7月1日C.R.達爾文與A.R.華萊士在倫敦林奈學會上宣讀了關於進化論的論文。後人稱他們的自然選擇學說為達爾文·華萊士學說。
主要觀點

進化論
第一,物種是可變的,生物是進化的。當時絕大部分讀了《物種起源》的生物學家都很快地接受了這個事實,進化論從此取代神創論,成為生物學研究的基石。即使是在當時,有關生物是否進化的辯論,也主要是在生物學家和基督教傳道士之間,而不是在生物學界內部進行的。
第二,自然選擇是生物進化的動力。生物都有繁殖過盛的傾向,而生存空間和食物是有限的,生物必須“為生存而鬥爭”。在同一種群中的個體存在著變異,那些具有能適應環境的有利變異的個體將存活下來,並繁殖後代,不具有有利變異的個體就被淘汰。如果自然條件的變化是有方向的,則在歷史過程中,經過長期的自然選擇,微小的變異就得到積累而成為顯著的變異。由此可能導致亞種和新種的形成。
理論缺陷
一般對進化論的批判在於認為其缺乏足夠的化石證據解釋不同物種之間的“過渡”,事1937年T.杜布尚斯基的《遺傳學與物種起源》一書提供解決方法。
理論上,若鳥類為爬行動物進化而來,從爬行動物至鳥類必然出現“過渡物種”,爬行動物為冷血動物,鳥類的體溫卻常維持比一般生物更高的溫度,中間的過渡物種如何平衡這樣的生理差異,還只能猜測,藉由到2015年為止,已發現大量的過渡物種化石,及孔夫子鳥和其他發現的化石證據顯示不但鳥類和恐龍曾經並存很長一段時間,且有部分恐龍是溫血動物,有種解釋是鳥類和恐龍的祖先是在更早的時間點就進化出溫血機制的蜥蜴(過渡物種),再藉由這種溫血蜥蜴進化出部分溫血的恐龍而後進化出鳥類。諸如此類的“進化先後”及“過渡物種”的問題有很多,且大部分沒有得到解決。主要原因在於大部分預言中的過渡物種都沒有找到完整化石,包括與人類祖先關係密切的南猿和古猿及古類人猿和直立行走的猿人之間的完整過渡化石。
根據模擬,生物進化中由“有機物到單細胞原核生物”和由“簡單的多細胞生物到複雜的進行了細胞分化的多細胞生物”兩個瓶頸難以被突破。而且,有一篇研究報告《生物沒有時間等待》指出,撇開這兩大問題不談。由單細胞生物開始以“基因的有利突變”方式進化至比靈長類低的生命形式需要近120億年的時間,所以此報告認為進化過程不可能是在地球上全部完成的,這個觀點因“現代綜合進化論”的遺傳應用的根本缺陷而產生,否定獲得性狀可以遺傳的“現代綜合進化論”無法解釋一致性持續突變及相關的進化速率問題。
由於達爾文的經典進化論具有極大的局限性,二十世紀的科學家對達爾文進化論進行了大刀闊斧的增加與修改,逐漸形成了現代進化論體系。
1865年奧地利植物學家G.J.孟德爾從豌豆的雜交實驗中得出了顆粒遺傳的正確結 論。他證明遺傳物質不融合,在繁殖傳代的過程中,可以發生分離和重新組合。
論。他證明遺傳物質不融合,在繁殖傳代的過程中,可以發生分離和重新組合。
進化論
20世紀初遺傳學建立,T.H.摩爾根等人進而建立了染色體遺傳學說,全面揭示了遺傳的基本規律。但把遺傳學說用在進化論上,會導向種族、血統決定論,所以當時大多數遺傳學家,都反對含有進化意義的達爾文自然選擇學說(摩爾根反對的是達爾文的性選擇學說Sexualselection)。
20世紀20~30年代首先由R.A.費希爾、S.賴特和J.B.S.霍爾丹等人將生物統計學與孟德爾的顆粒遺傳理論相結合,重新解釋了達爾文的自然選擇學說,形成了群體遺傳學。
以後C.C.切特韋里科夫、T.多布然斯基、J.赫胥黎、E.邁爾、F.J.阿亞拉、G.L.斯特賓斯、G.G.辛普森和J.W.瓦倫丁等人又根據染色體遺傳學說、群體遺傳學、物種的概念以及古生物學和分子生物學的許多學科知識,發展了達爾文學說,建立了現代綜合進化論。
現代進化理論認為,進化是生物種群中實現的,而突變、選擇、和隔離是生物進化和物種形成過程中的三個基本環節。現代綜合進化論徹底否定獲得性狀的遺傳,強調進化的漸進性,認為進化是群體而不是個體的現象,並重新肯定了自然選擇的壓倒一切的重要性,繼承和發展了達爾文進化學說。
根據新的觀察發現,遺傳變異來源於DNA損傷,而DNA損傷修復中有一種直接跨過DNA損傷進行合成的特殊的DNA聚合酶被稱為跨損傷合成聚合酶,如果修復中DNA遺傳信息產生變異,則必然與此酶蛋白遺傳信息相關。
遺傳與變異是矛與盾的對立面,遺傳存在於DNA複製,屬於遺傳複製經驗總結的中心法則不能越界論定變異方面的如DNA損傷中蛋白質信息逆傳遞問題,“現代綜合進化論”對獲得性狀遺傳的“徹底否定”不成立。與環境適應性相關的進化速率的漸進與躍變、進化的群體性等各方面,更符合生物史地質史的是獲得性狀可以遺傳的拉馬克進化觀。
最近,有學者對現代各種進化論進行了綜合,提出了進化“四因說”,認為基因是遺傳的質料,基因組儲存了生命形成的原則(形式),個體在初生動因(太陽光能)和次生動因(遺傳的、生理的、生態的)的聯合驅動下,通過求生(一種次生的目的性),推動著種族的延綿與分化。進化是以無數個體生命為載體的一種生命運動,它也是一種物質運動形式,雖然比物體的物理學運動要複雜的多。生命存在的實體—個體是質料、形式、動因和目的的統一,其自身是多種生命層次運動的複合,也是諸多類型運動的複合,這些進一步形成了種族的生命運動—演化或進化。
進化論是指研究生物史中生命擴大適應範圍,增加生存餘地的發展狀況的科學結論(反進化觀點認為生物史無進化無方向無結論的人因此宣傳另一個不含達爾文進化思想的理論並命名為“演化論”)。隨著進化論的發展,產生了現代綜合進化論,而現代進化學絕大部分以查爾斯·羅伯特·達爾文的進化論為指導,埃爾溫·薛定諤的《生命是什麼》為主體方向,進化論已為當代生物學的核心思想之一。其進化論有三大經典證據:比較解剖學、古生物學和胚胎髮育重演律。

進化論
種群的基因頻率若保持相對穩定,則該種群的基因型也保持穩定。但在自然界中種群基因頻率的改變是不可避免的,於是基因型也逐漸變化。
2.突變為生物進化提供材料。突變引起的基因頻率的改變是普遍存在的。誠然,突變發生的自然頻率是相當低的,例如改變染色體數目(降低後代繁殖率)的染色體平衡易位(羅伯遜易位或羅氏易位)在人類的發生律只有1/500~1/1000,種群是由大量的個體組成,每個個體具有成千上萬基因,這樣,每一代都會產生大量的變異。
突變的結果可形成多種多樣的基因型,使種群出現大量可遺傳變異。這些變異是隨機性的,不定向的,能為生物進化提供原料。但突變大多有害,這是理論難以解釋的。
3.自然選擇主導著進化的方向。突變的方向是不確定性的,一旦產生,就在自然界中受到選擇的作用。自然選擇不斷淘汰不適應環境的類型,從而定向地改變種群中的基因頻率向適應環境的方向演化。
自然選擇不斷地調整著生物與環境的關係,定向地改變種群的基因頻率。那麼,自然界又如何將改變了的基因頻率在種群中相對固定下來,進而形成新的適應於環境的生物類型呢?
4.隔離是物種形成的必要條件。隔離使不同物種之間停止了基因交流,一個種群中所發生的突變不會擴散到另一個種群中去,使不同的種群朝不同的方向演化。隔離一般分為地理隔離和生殖隔離兩類。地理隔離是由於某些地理障礙而發生的。大河、大山、沙漠、海峽和遠距離都能將種群阻隔開來,使他們之間彼此不能往來接觸,失去了交配的機會。長期的地理隔離使兩個種群分別接觸不同的環境,各自積累了變異。另一方面,長期的地理隔離使相互分開的種群斷絕了基因交流,結果導致了生殖隔離。生殖隔離是指進行有性生殖的生物彼此之間不能雜交或雜交不育。生殖隔離又可分為受精前的生殖隔離和受精后的生殖隔離。生殖隔離一旦形成,原來的一個物種的種群就變成兩個物種的種群了。

進化論
缺少了進化中有幾種類型的基因驅動描述,包括基於歸巢內切酶的基因驅動、性連鎖減數分裂驅動、medea基因驅動和顯性不足基因驅動。他們從不同的屬性上描述了這些基因驅動,包括擴散速率、物種特異性、適合度代價(fitnesscost)、抵抗易感性、可移除性和可逆性。這些都是進化生物學的數學描述
達爾文過分強調了生物進化的漸變性;他深信“自然界無跳躍”,用“中間類型絕滅”和“化石記錄不全”來解釋古生物資料所顯示的跳躍性進化。傳統進化論的這種觀點正越來越受到間斷平衡論者和新災變論者的猛烈批評。當時的生物學家對接受這一點猶豫不決,因為自然選擇學說在發表時存在著三大困難。
第一:缺少過渡型化石。按照自然選擇學說,生物進化是一個在環境的選擇下,逐漸地發生改變的過程,因此在舊種和新種之間,在舊類和新類之間,應該存在過渡形態,而這隻能在化石中尋找。在當時已發現的化石標本中,找不到一具可視為過渡型的。達爾文認為這是由於化石記錄不完全,並相信進一步的尋找將會發現一些過渡型化石。確實,在《物種起源》發表兩年後,從爬行類到鳥類的過渡型始祖鳥出土了,以後各種各樣的過渡型化石紛紛被發現,最著名的莫過於從猿到人的猿人化石。如今被稱為過渡型的化石已有上千種,與已知的幾百萬種化石相比,仍然顯得非常稀少。這有兩方面的原因。一方面,生物化石都是偶然形成的,因此化石記錄必然非常不完全;另一方面,按照現代進化論體系所提出的“間斷平衡”理論,生物在進化時,往往是在很長時間的穩定之後,在短時間內完成向新種的進化,因此過渡形態更加難以形成化石。
第二:地球的年齡問題。既然自然選擇學說認為生物進化是一個逐漸改變的過程,它就需要無比漫長的時間。達爾文認為這個過程至少需要十幾億年。但是當時物理學界的泰斗威廉·湯姆遜(即開爾文勛爵,一個神創論者)用熱力學的方法證明地球只有一億年的歷史,而只有最近的最多兩千萬年地球才冷卻到能夠讓生命生存。對於物理學家的挑戰,達爾文無法反擊,只能說“我確信有一天世界將被發現比湯姆遜所計算而得的還要古老”。在現代物理學支持此下,如今的地質學界鈾鉛測年法計算出地球有四十幾億年的歷史,而至少在三十億年前生命就已誕生。在當時,在地球的年齡問題上,人們顯然更傾向於相信物理學權威。
第三:無法解釋自然選擇。達爾文找不到一個合理的遺傳機理來解釋自然選擇。當時的生物學界普遍相信所謂“融合遺傳”:父方和母方的性狀融合在一起遺傳給子代。這似乎是很顯然的,白人和黑人結婚生的子女的膚色總是介於黑白之間。湯姆遜的學生、蘇格蘭工程師簡金(F.Jenkin)據此指出:一個優良的變異會很快地被眾多劣等的變異融合、稀釋掉,而無法像自然選擇學說所說的那樣在後代保存、擴散開來,就像一個白人到一個非洲黑人部落結婚生子,幾代以後他的後代就會完全變成了黑人。達爾文雖然從動植物培養中知道一個優良的性狀是可以被保留下來的,但是他沒有一套合理的遺傳理論來反駁簡金。達爾文被迫做出讓步,承認用進廢退的拉馬克主義也是成立的,可以用來補充自然選擇學說。
第四:達爾文晚年看法。
1915年在一份美國報章第一次發表的作者不詳的文章TheLadyHopeStory,作者提出達爾文老年在病榻時再度信奉基督教,卻被達爾文的子孫及歷史學家否認。他的女兒表示,達爾文未再信奉基督教,他死前最後一句說話是對妻子說“記著,你一直是一位多麼好的太太。”縱然如此,很多反進化論者仍然會用《TheLadyHopeStory》里“達爾文死前再信奉基督教”的陳述來指出達爾文自己否定了進化論,藉此宣傳反進化論。
科學和各大宗教衝突自西方啟蒙時代開始已經爭鬧不休,西方中世紀教會擁有極大的權力,政教是合一的。教會和皇權操控社會一切活動和個人生活細節。學術研究更加不能夠逾越教會正統信仰的教導。基督教教會,包括天主教和宗教改革產生的宗派,相信人是神按神形象創造,人在一切創造物是至為尊貴的。因此,當時的人都是按照聖經的教導解釋所有對自然現象的理解,教會傳統教導是無誤的,天文學﹑地理學﹑生物學、哲學等學科,都是教會權威,不可以挑戰,學術的責任是發展知識服務教會﹑去支持教會和聖經無誤的教導,這一點於啟蒙運動和宗教改革后二者得以分離。
進化論給人文領域帶來了一場顛覆性的革命,在達爾文之前,人們普遍認為人是萬物之靈,並非自然界的一部分,而是超越了自然。例如猶太教及基督教神學把人看成上帝根據自己的形像創造出來的特殊作品,在世界萬物中只有人才被賦予了靈魂,世界萬物都是被創造出來為人服務的,人與其他動物存在不可逾越的鴻溝。
達爾文指出,人類是生物進化過程中的偶然產物,大自然的產物。今天的一切生物都是人類的親屬,人類與其他生物特別是與類人猿並無本質的區別,我們認為人類特有的屬性——例如智力、道德觀等精神因素——都可在其他動物中找到雛形,也必定有其自然的起源。進化論推翻了人類自以為與其它動物不同的地位,人類和猿猴分享同一個祖先,這是難以令人接受的。對於信奉基督教的人士而言是無法接受的,這種思維直到世界性政教分離后才得以理性化。
西方國家一些宗教團體認為演化論與創世理論有衝突,某些地區的學校甚至不開設演化論的課程,而一些地區開設演化論課程是與宗教團體妥協的結果。現代社會以美國為例,美國弗羅里達州規定必須在公立學校教授演化論,2005年法院以政教分離原則判進化論是科學理論,不得與宗教信仰混為一談。
2017年6月,土耳其推進教改,繼中小學教材刪除達爾文進化論、壓縮國父凱末爾相關內容之後,土耳其教育部門宣布,停止高中階段教學達爾文的生物進化論,稱進化論是“有爭議的且難以理解”,因為有穆斯林背景的學生認為世界萬物是由真主安拉創造的。學校使用的標準生物教材中“生命的起源和進化論”這一章節將被刪除。只有高等教育階段的學生才能接觸這些材料。
有些有宗教信仰的科學家認為“演化論只是闡明了神是如何創造生物的,並非否定創造論”,現代社會多數教會(包括天主教會)認同此觀點。但神導進化論與創世紀的前幾章相矛盾,且無法解釋人的罪性從何而來,也就不能說明耶穌為何要被釘死在十字架上。
2005年,在美國一封由1萬多名基督教牧師簽名的牧師公開信說:“……我們相信進化論是科學的基本事實,它經受住了嚴格的檢驗,人類許多豐富的知識和成就都建築在這個理論之上。拒絕這個事實,或者認為它只不過是‘許多假說中的一種’,這是對科學無知的蓄意容忍,而且將把這種無知傳遞給我們的後代。我們相信,能夠進行批判性思維的大腦就是上帝給予我們的珍貴禮物之一,拒絕充分使用這個禮物就是拒絕造物主的意願……我們敦促學校董事會成員肯定進化論教學是人類知識的重要部分,並因此維護科學課程的完整性。我們要求,把科學作為科學來對待,把宗教作為宗教來對待,這是兩種不同形式的相互補充的真理。”
是否存在這篇公開信這個事實需要考證。
在進化論中討論智能設計的主因是因為智能設計的主要目的在於打擊進化論及取代進化論在生物課上出現,並以不違法的原則傳授神創論,進而成為政治,宗教,及科學的核心,而非為了對科學進步有所貢獻。
這個理論除了攻擊進化論的部分爭議(大部分的"爭議"在生物學家看來已經沒有爭議)外,本身並沒有具有邏輯思維的論點、直接證據或是任何發表於科學期刊的文獻可以做為支持智能設計的理論基礎。
它相信世界上所有的生物都來自一個智能設計者(改自上帝),在一瞬間的創造,且這些生物除了為同一創造者造出之外毫無共通之處,即使之間具有差異也不可能改變物種。其理論的基礎是聖經的創世紀(參考奇茲米勒訴多佛學區案)。
某些基督教團體認為智能設計是同等重要的科學理論,甚至比現有進化論對生命起源問題的解釋更加合理,可后其被否定。
2005年奇茲米勒訴多佛學區案一案。在這案例中原告方提出了證據證明當初智能設計的原稿只是份講解聖經創世紀用的文章,於1987年由作者在Edwardsv.Aguillard一案后,將"創造者"改為"智能因子"的手法舊瓶換新葯的神創論。
在《貓熊與人》一書(智能設計的主要文獻之一)中曾出現過一個“Cdesignproponensist(“創”設計者,錯字)”,實為將“Creationist(創造者)”改為“designproponensist(設計者)”時編輯失誤造成。
進化定義
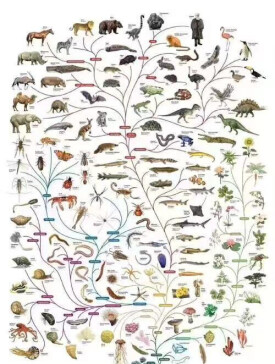
從進化的觀點看,同時生存的不同生物種在時間的向度上可以回溯到一個共同祖先。因此,按照祖裔關係可以將現時生存的和曾經生存過的生物相互聯結起來,這種表示祖裔關係的生物進化系統稱為種系發生。生物的種系發生可以形象地表示為一棵樹:如果從樹根到樹頂代表時間向度,主幹代表共同祖先,大小分枝代表相互關聯的進化線系,這就構成所謂種系發生樹或進化樹。所謂進化型式就是進化在時間與空間上的特徵,也就是種系發生的特徵,具體表現在進化樹的形態上:枝幹的延續和分枝方式、樹榦的傾斜方向和在空間上的配置、樹榦的中斷等等,它代表著種系發生中線系進化、種形成、絕滅等方面的特徵。
在譜系進化中有兩種性質不同的進化改變。一是形態結構及其功能由簡單、相對不完善到複雜和相對完善的前進性(進步的)改變,稱之為前進進化;前進進化的結果是造成生物的等級從低級到高級。另一種進化改變是線系分枝,叫做分枝進化;分枝進化的結果是產生新的分類單元和生物歧異度的增長。廣義的前進進化包括除分枝進化以外的各種進化改變,既包括前進的(進步的)進化改變,也包括非前進的甚至退化的改變。既無前進進化,又無分枝進化的情況稱為停滯進化,如活化石之類的情況。
適應輻射
在相對較短的地質時間內,從一個線系分枝出許多歧異的分類單元,叫做輻射。由於輻射分枝通常是向不同的方向適應進化的,所以又稱為適應輻射。
不同的線系各自獨立地進化出相似的特徵叫做“趨同進化”,簡稱“趨同進化”。形態結構的進化趨同往往是由於功能的相似,而功能的相似又往往是由於適應於相似的環境。
平行進化
(簡稱平行)是指兩個或多個有共同祖先的線系,在其祖先遺傳的基礎上分別獨立進化出相似的特徵。通常平行與趨同不易區分,一般說來平行進化既涉及同功又涉及同源; 假若後裔間的相似程度大於各自的祖先之間的相似程度則可稱為趨同,若後裔之間的相似程度與其祖先之間的相似程度差不多則可稱為平行。大進化的模式①漸變模式。認為形態進化速度多少是恆定的、勻速的,形態進化是逐漸的;形態改變主要是線系進化造成的,大多發生在種的生存期間,與種形成無關,種形成(線系分枝)本身只是增加新的進化方向。②間斷模式。認為形態進化速度是不恆定、非勻速的,快速地“跳躍”與長期的停頓相交替,即在種形成期間進化加速,種形成后保持相對的穩定。進化的形態改變與種形成相關聯,即大部分形態改變是發生在相對較短的種形成時期,在可能長達數百萬年的種生存期內不會發生顯著的形態改變。兩種模式各有一些有利於自己的證據,這些證據本身還需進一步研究才能證實。
假若後裔間的相似程度大於各自的祖先之間的相似程度則可稱為趨同,若後裔之間的相似程度與其祖先之間的相似程度差不多則可稱為平行。大進化的模式①漸變模式。認為形態進化速度多少是恆定的、勻速的,形態進化是逐漸的;形態改變主要是線系進化造成的,大多發生在種的生存期間,與種形成無關,種形成(線系分枝)本身只是增加新的進化方向。②間斷模式。認為形態進化速度是不恆定、非勻速的,快速地“跳躍”與長期的停頓相交替,即在種形成期間進化加速,種形成后保持相對的穩定。進化的形態改變與種形成相關聯,即大部分形態改變是發生在相對較短的種形成時期,在可能長達數百萬年的種生存期內不會發生顯著的形態改變。兩種模式各有一些有利於自己的證據,這些證據本身還需進一步研究才能證實。
進化論
進化速率
單位時間內生物進化改變數。衡量進化速率必須確定兩個尺度:時間尺度和進化改變數的尺度。時間尺度有兩種,即絕對地質時間和相對地質時間,一般應使用絕對時間尺度;只在不得已時,才使用相對時間尺度。
進化趨勢
從長的時間尺度來看生物進化呈某種方向性,但這並非說自然界存在著既定的進化軌道,這裡說的方向性是統計學的趨向。因此,進化趨勢是任何因素(包括隨機的非隨機的因素)所造成的進化的統計學趨勢。
1)小進化趨勢:是指在生物種的自然群體之內,自然選擇作用所造成的表型進化改變的趨向性。這種進化趨勢通常是適應局部環境的結果,進化改變發生在線系之內,故也可稱之為線系進化趨勢,表現為線系在時間-進化改變坐標系內的傾斜。
2)大進化趨勢:是指在種系發生中線系分枝的主體的趨向性。大進化趨勢發生在種以上的分類單元的譜系進化中,所以又可以稱為譜系進化趨勢,表現為譜系中的偏斜。為解釋大進化趨勢發生的原因,不少學者提出過種種假說,但由於缺乏證據,能夠成立或有說服力的還很少見。進化是生物逐漸演變向前發展的過程。在這個過程中,生物由低級發展到高級,由簡單發展到複雜。今天地球上的各種生物,極少是和遠古時代的祖先一模一樣的。同樣,在未來,各種生物又會和今天不同,這就是進化的結果。進化的過程是極其緩慢的,要經過長期的自然選擇逐漸地演化。除了由低等進化到高等外,生物的種類也不斷地增多。今天的物種遠比5億年前的物種多得多,今後還會不斷地增加。
進化方向
拉馬克主義信奉終因論,認為進化實質上就是從低等走向高等,從原始走向高級,從簡單走向複雜,從不完美走向完美。如果沒有這種內在力量,如何解釋生命從最簡單的細菌逐漸進化出美麗的蘭花、參天的大樹、迷人的蝴蝶和智慧的人類呢(邁爾2010)?在這一理論框架中,進化是進步式的,即生物進化的目的就是走向更加完美,人類不是一個普通的進化分枝,而是生物進化的頂點,獲得性遺傳中所隱含的生物努力可以得到回報—生物在一代發生的變化可以遺傳下去。
而達爾文則堅決反對這種神秘力量的存在。按照達爾文的進化論,自然界中不存在目的,不具方向性,人類僅是進化中的一個普通分支。達爾文對昆蟲與植物傳粉之間的關係很感興趣,在這裡,動物中種類最多的昆蟲和植物中種類最多的顯花植物之間的協同進化的確充滿了隨機性,並未呈現出明確的方向性。
從拉馬克主義衍生出來的直生論者(主要是一些古生物學家)信奉內在的定向的力量。伯格LS提出了循規進化論(nomogenebis)—生物的進化是個有規則的過程,隨機變異和環境的影響都不能使這個過程發生改變,他認為多數進化是進步的,使生物不斷完善,而且在進化中特意的性狀規則地出現。奧斯本HF提出了芒狀發生說(aristogenesis),認為在綱以上的階元,譜系的發生按照共同由來的原則進行,在綱以下的階元,則通過“適應輻射”產生許多分支,之後,這些分支的進化則成直線狀,不再分化,這樣整個譜系呈芒狀(Bowler1983,見田洺1998)。
謝平認為,進化既不完全隨機,也不完全定向,在隨機中有一定的方向性,在方向中有一些隨機性,這本質上源於生命的一種拼接與堆砌式的物種創造方式。舉個通俗的例子,人再怎麼隨機變異,也不可能突變為昆蟲,昆蟲無論如何也不可能隨機突變成人。一個類群的演化歷史已經大致決定了它繼續分化的方向,這在複雜的高等動物中最為明顯,即物種的結構越複雜、特化程度越高(可能源於日益加劇的種間競爭與日趨精細的生態位分化),這種變化的方向可能越狹窄,因為它能自由調整的餘地就越小,甚至走向滅絕之路,這並不是設計的天意,而是因為過分大型化、複雜化和特化的身體構造對環境寬幅波動的適應性變得越來越差的一個可悲的結局。恐龍或許就是這樣的例子,而為了避免一味大型化而向社會化或群體性動物的發展也許能開闢一條成功之道,這似乎詮釋了后恐龍時代為何哺乳動物得以興盛。
達爾文進化論認為生物進化是勻速的,在解釋一些問題上遇到了困難,,比如2000萬年間就發生了寒武紀生命大爆發。寒武紀生命大爆發被稱為古生物學和地質學上的一大懸案──寒武紀生命大爆發,自達爾文以來就一直困擾著進化論等學術界。大約6億年前,在地質學上稱做寒武紀的開始,絕大多數無脊椎動物門在只2000多萬年時間內出現了。這種幾乎是“同時”地、“突然”地在2000多萬年時間內出現在寒武紀地層中門類眾多的無脊椎動物化石(節肢動物、軟體動物、腕足動物和環節動物等),而在寒武紀之前更為古老的地層中長期以來卻找不到動物化石的現象,被古生物學家稱作“寒武紀生命大爆發”,簡稱“寒武爆發”。
寒武爆發代表了生物進化過程中的真實事件,科學家從物理環境和生態環境的變化兩個方面來解釋這一現象。
“間斷平衡”理論認為,生物的進化不像達爾文及新達爾文主義者所強調的那樣是一個緩慢的連續漸變積累過程,而是長期的穩定(甚至不變)與短暫的劇變交替的過程,從而在地質記錄中留下許多空缺。在間斷平衡學說的支持者們看來,大進化有著與微進化不同的機制,而這種大進化機制,不是自然選擇,而是其他因素所致—譬如“發育制約”—胚胎髮育的模式(藍圖)一旦建立起來,就有了一種內在的連貫性,難以通過突變逐漸加以改變,生物將沿著固定的途徑發育、生長,使物種長期保持穩定,新的遺傳變異由於不能與已有的發育模式相容,因此不可能出現或保留下來。
作為20世紀最驚人的科學發現之一的中國雲南澄江動物群,它是世界上目前所發現的最古老、保存最為完整的帶殼後生動物群。
該動物群是中國青年古生物學家侯先光1984年在雲南澄江縣帽天山首先發現的。這是一個內容十分豐富、保存非常完美,距今約5.7億年的化石群,其成員包括水母狀生物、三葉蟲、具附肢的非三葉的節肢動物、金臂蟲、蠕形動物、海綿動物、內肛動物、環節動物、無絞綱腕足動物、軟舌螺類、開腔骨類,以及藻類等,甚至還有屬於低等脊索動物或半索動物(如著名的雲南蟲)等。由於許多動物的軟組織保存完好,為研究早期無脊椎動物的形態結構、生活方式、生態環境等提供了極好的材料,同時也成為了探索地球上大殼後生動物爆發事件的重要窗口。
地球膨裂說認為,因為地球寒武紀溫度的不斷升高,非常適合生物繁衍,進化速度迅速增加,所以發生了寒武紀生命大爆發。因此有人認為寒武紀生命大爆發不但不是反對進化論的有力證據,反而是進化論的有力證據。
1859年,達爾文發表了經典巨著《物種起源》,對生命起源和演化這一極其漫長而又紛繁複雜的過程提出了“進化論”的科學解釋。
美判“智能設計論”違憲禁上課堂。
科學本身並不否定自己的錯誤和不足。有人認為澄江生物群的發現說明了生物的進化並非總是漸進的,而是漸進與躍進並存的過程。但躍進理論缺乏依據,無法解釋其原因和具體過程。
眾多的神創論引用的寒武紀大爆發,其實確實只是一個數百萬年的過程,並不漫長,而雖然最近的化石證據表明,在之寒武紀前,存在著大量多細胞生物,但不能這說明它們與之後的複雜生物有親緣關係,所以進化論不一定是正確的,只是一個假說,生物起源理論還需要不斷完善,科學絕對不會局限於此。
異質系克隆:以進化論模型構建合胞滋養層分析構建的內源性病毒進化樹。選擇適配的低排異母體
蛋白質體外定向進化:2018年諾貝爾化學獎,過催化化學反應方式定向進化產生的蛋白質有著廣泛的應用領域,包括生物燃料、藥品等等
基本信息
- 中文名
- 進化論
- 外文名
- Theory of evolution
- 拼音
- jìn huà lùn
- 應用學科
- 生物學
- 別名
- 演化論、天演論
- 發展
- 現代綜合進化論
- 適用領域
- 進化生物學
- 提出者
- 布豐和拉馬克最早提出,達爾文為公認的集大成者