細胞起源
細胞起源
一般認為,地球上生命的演化或進化歷史包含了從原始的前細胞進化出原核細胞進而到真核細胞的複雜過程。但是,細胞到底是如何起源的依然是現代生命科學中依然未解的最大謎團之一。原核細胞的成型也就標誌著原始地球上生命的正式誕生,而真核細胞的誕生伴隨了地球生命的空前繁榮。
“細胞”一詞原指有細胞核的真核細胞。恩格斯在《反杜林論》、《自然辯證法》等著作中所指的即這一意義的細胞。50年代以來“細”的含義有了擴大,把原核細胞即細菌、藍藻等無核生物也包括進來,因而細胞起源和生命起源的界限便不易區分了。細胞的起源包括原核細胞的起源和真核細胞的起源問題。
在生命進化歷史上,細胞是怎樣從原始的前細胞的生命形式進化而來的?從化石記錄可以確定原核生物在距今30~35億年前即已出現,先於真核生物。在此以前,應存在漫長的化學進化過程:地球原始大氣中的N,NH,CH,CO在高溫、強紫外線或放電條件下形成含碳化合物和生物大分子的前體物質(如氨基酸、核酸鹼基等);然後通過聚合形成複雜的生物大分子並且在原始的海水中出現大分子的複合物,稱為團聚體或類蛋白小體;隨著遺傳密碼(核酸)的出現,才有可能出現自身繁殖的細胞。
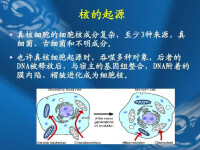
圖1
最近,有學者提出了細胞起源的光合偶聯假設(photosynthesis-coupling hypothesis)。在一定意義上來說,生命的起源始於有機物質的個性化體系的建立,或者說,沒有個性便不會有真正生命的誕生。試問,如果有機分子只是飄忽無定地在“湯”中蕩漾,怎會出現現在的生命?哪種生命不以個體形式而存在呢?因此,能導致獨立生命系統形成的簡單的質膜結構的出現(不論通過何種機制或過程)必定是細胞進化的最關鍵的一步。不僅需要前細胞體整體的獨立性,一些重要生命過程(特別是光合作用)也需要細胞內的獨立性,即需要內膜系統,看看藍細菌的專門的光合機構—類囊體就是如此。
在具備簡單膜結構的前細胞體中,光合作用指向的選擇過程促進主要生命物質的功能分化—脂質構建細胞膜、蛋白質主要扮演催化者、DNA儲存遺傳信息、RNA構成DNA與蛋白質之間的橋樑、糖類暫時性儲存光合作用轉化而來的化學能……。這不完全只是隨機過程(雖然在很大程度上是),它更是具有目的性指向(最有效的太陽光能利用)的化學性與生命性的巧妙融合,它為生化反應的秩序化與生命過程的程序化鋪墊了基石。
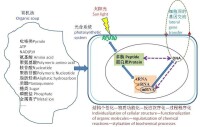
圖2細胞起源的光合偶聯假說
生命的另一個本質特徵就是將遺傳物質指揮生命構建過程程序化,即將生命構建過程信息化地儲存於DNA之中,這是個性化的生命獲得自我繁殖特性必須邁出的重要一步。最後是細胞分裂機制的逐漸形成,這得益於細胞膜的半通透性(小分子養分的進入與大分子碳水化合物的堆積),以及二者之間的矛盾及其必然的結果—細胞破裂過程的無數次重複。精確而周期化的細胞分裂才能使穩定的遺傳成為了可能,這時像我們在現存生物中能所見到的細胞才成功地誕生了,細胞運行機制才得以程序化了。
細胞核起源依然是未解之謎,迄今為止提出的主要學說有:共營模型(syntrophic model)、自演化模型(autogenous model)、病毒性真核生物起源模型(viral eukaryogenesis model)、外膜假說(exomembrane hypothesis)、壓縮和結構化假說(packing and structurization hypothesis),等等。目前設想的關於“核”起源的流行情景是所謂的內共生途徑。
支持線粒體和葉綠體共生起源假說的主要論據,是它們都是能自身複製的半自主結構,都含有自己的DNA和獨立的蛋白質合成裝置,包括結構上與真核細胞的細胞質核糖核蛋白體不同而與真細菌類相似的核糖核蛋白體。與原核生物相似,葉綠體的DNA以及動物細胞線粒體的DNA均呈閉環形。此外,線粒體、葉綠體的蛋白質合成裝置在功能上還存在與真細菌類相似的特點。例如,氯黴素能抑制線粒體、葉綠體和真細菌類之核糖核蛋白體的蛋白質合成,但不能抑制真核細胞細胞質核糖核蛋白體的蛋白質合成。反之,放線菌酮能抑制真核細胞細胞質核糖核蛋白體的蛋白質合成,而不影響真細菌類或線粒體、葉綠體的蛋白質合成。線粒體和葉綠體在結構和功能上與真細菌類的這些相似之處,均提示這兩種細胞器可能是從細胞內共生的真細菌類演變來的。在這一轉變過程中,原有的一些必要基因轉移到宿主細胞核內,一些無用的基因則逐漸喪失,結果成為在代謝上與宿主互補的半自立的結構。至於中心粒是否含有DNA,目前尚無定論,不過它顯然是能自身複製的。但是,馬古利斯(Lynn Margulis)的假說並未涉及細胞核的起源。一般認為原始的細胞核是由質膜內褶產生的內質網包圍了原核生物的染色質結構而形成的,多少近似於現今的渦鞭毛蟲類的核。
假想的最原始的單細胞真核生物葉綠體和線粒體的內共生起源假說有一個理論前提,就是必須設想最原始的單細胞真核生物或前真核生物已經具有進行吞噬活動的能力,否則不可能攫取到胞內共生物。
已知有些原細菌具有近似肌動蛋白的蛋白質。因此,可以設想由古代原細菌起源的前真核生物可能已有了肌動蛋白,而這就為它們進行胞質環流、變形運動和吞噬活動創造了條件。
有利於這種設想的一個論據是對各類低等真核生物的5S rRNA與原細菌類的5S rRNA一級和二級結構的比較,發現在某幾點上同原細菌類最接近的是變形蟲。這意味著變形蟲類可能比其他各類低等真核生物(如綠藻、真菌等)都更為原始。
考慮到渦鞭毛蟲類的細胞核所表現出來的原始特性,可以設想最初發生的最原始的單細胞真核生物或許是一種具有渦鞭毛蟲式的細胞核的變形蟲。
基本信息
- 中文名
- 細胞起源
- 外文名
- Origin of cells
- 定義
- 原核細胞的起源和真核細胞的起源
- 類型
- 科學術語