間隙連接
連接子進行的細胞間連接
間隙連接也可以稱為連接或黃斑通信。 (雖然大多數神經組織沒有間隙連接,但是當在神經元或神經中發現時,它也可以被稱為電突觸,如牙髓中的神經細胞。)雖然ephapse與間隙連接有一些相似之處,但是通過現代定義,兩個是不同的。
間隙連接是多種動物細胞類型之間的特殊細胞間連接。它們直接連接兩個細胞的細胞質,使各種分子,離子和電脈衝直接通過細胞之間的調節門。
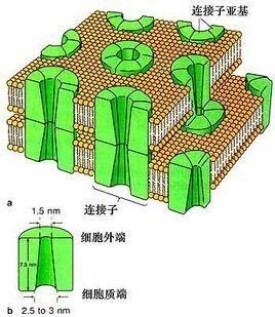
一個間隙連接通道由兩個連接子(或半通道)組成,它們跨越細胞間隙連接。間隙連接類似於連接植物細胞的胞間連絲。
除了成人完全發育的骨骼肌和移動細胞類型(如精子或紅細胞)之外,幾乎在身體的所有組織中都存在間隙連接。然而,在諸如海綿和粘液黴菌之類的較簡單的生物體中沒有發現間隙連接。
在脊椎動物中,間隙連接半通道主要是連接 蛋白的同源或異源 六聚體。無脊椎動物間隙連接包含來自innexin 家族的 蛋白質。 Innexin與連接蛋白沒有顯著的序列同源性。雖然序列與連接蛋白的序列不同,但innexins與連接蛋白相似,表明innexins以與連接蛋白相同的方式在體內形成間隙連接。最近表徵的pannexin家族最初被認為形成細胞間通道(具有與innexins相似的氨基酸序列),實際上起到單膜的作用與細胞外環境相通的通道,已被證明可以通過鈣和ATP。
在間隙連接處,細胞間隙在2到4nm之間並且每個細胞膜中的單位連接子彼此對齊。
由兩個相同的半通道形成的間隙連接通道稱為同型,而具有不同半通道的那些是異型的。反過來,均勻連接蛋白組成的半通道被稱為同源,而具有不同連接蛋白的那些是異聚的。通道組成被認為影響間隙連接通道的功能。
在對innexins和pannexin進行了充分表徵之前,基於基因定位和序列相似性,將編碼連接蛋白間隙連接通道的基因分為三組之一:A,B和C(例如,GJA1 ,GJC1 )。然而,連接蛋白基因不直接編碼間隙連接通道的表達; 基因只能產生構成間隙連接通道的蛋白質。
2.一種連接蛋白具有四個跨膜 結構域
3.6 Connexins創建一個Connexon(半通道)。當不同的連接蛋白連接在一起形成一個連接子時,它被稱為異聚連接子
4.跨細胞膜連接在一起的兩個半通道包括間隙連接通道。當兩個相同的連接子聚集在一起形成間隙連接通道時,它被稱為同型GJ通道。當一個同聚連接子和一個異聚連接子聚集在一起時,它被稱為異型間隙連接通道。當兩個異聚連接子連接時,它也稱為異型間隙連接通道。
5.幾個間隙連接通道(數百個)組裝在稱為間隙連接斑塊的大分子複合物中。
1.允許細胞之間的直接電通信,儘管不同的連接蛋白亞基可以賦予不同的單通道電導,從約30pS到500pS。
2.允許細胞之間的化學通信,通過傳輸小的第二信使,如肌醇三磷酸( IP )和鈣( Ca ),雖然不同的連接蛋白亞基可以賦予特定小分子不同的選擇性。
3.一般而言,允許小於485 道爾頓(通過無脊椎間隙連接 1,1 1,100道爾頓)的分子的跨膜運動,儘管不同的連接蛋白亞基可以賦予不同的孔徑和不同的電荷選擇性。大的生物分子,例如核酸和蛋白質,通過間隙連接連接蛋白通道被排除在細胞之間的細胞質轉移中。
4.確保通過間隙連接的分子和電流不會泄漏到細胞間隙中。
迄今為止,五種不同的功能歸因於間隙連接蛋白:
1.細胞之間的電和代謝耦合
2.通過半通道進行電和代謝交換
3.腫瘤抑制基因( Cx43 , Cx32和Cx36 )
4.粘附功能獨立於導電間隙連接通道(新皮層中的神經遷移)
5.羧基末端在信號傳導細胞質通路中的作用(Cx43)
已經在細胞彼此接觸的各種動物器官和組織中觀察到間隙連接。從20世紀50年代到70年代,他們在小龍蝦神經,大鼠胰腺,肝臟,腎上腺皮質,附睾,十二指腸,肌肉,水蚤 肝臟, Hydra肌肉,猴子視網膜,兔角膜,魚胚盤,青蛙胚胎,兔子卵巢,再聚集細胞,蟑螂血細胞膠囊,兔皮,小雞胚胎,朗格漢斯人類胰島,金魚和倉鼠壓力感測聲前庭受體,七鰓鰻和被囊心臟,大鼠曲細精管,子宮肌層,眼鏡片和頭足類消化上皮。自20世紀70年代以來,在幾乎所有相互接觸的動物細胞中都發現了間隙連接。到20世紀90年代,共聚焦顯微鏡等新技術可以更快速地檢測大面積組織。自20世紀70年代以來,甚至傳統上被認為可能具有分離的細胞(例如骨)的組織顯示細胞仍然與間隙連接相連,但是很有效。間隙連接似乎存在於所有動物器官和組織中,除了通常不與相鄰細胞接觸的細胞外,發現其他例外將是有趣的。成人骨骼肌是一個可能的例外。可以認為,如果存在於骨骼肌中,間隙連接可能在構成肌肉的細胞之間以任意方式傳播收縮。至少在某些情況下,可能不是如其他具有間隙連接的肌肉類型所示的情況。通過分析癌症 或衰老過程可以指示間隙連接減少或缺失的結果。
可以看到間隙連接在最簡單的水平上起作用,作為電流,小分子和離子的直接細胞到細胞途徑。如下所述,對這種通信的控制允許對多細胞生物的複雜下游效應。
在20世紀80年代,已經研究了間隙連接通信的更微妙但同樣重要的作用。發現通過將抗連接蛋白抗體添加到胚胎細胞中可以破壞間隙連接通訊。具有阻塞間隙連接區域的胚胎未能正常發育。抗體阻斷間隙連接的機制尚不清楚,但進行了系統研究以闡明其機制。這些研究的細化表明,間隙連接似乎是細胞極性發展的關鍵和動物的左/右對稱/不對稱。雖然確定身體器官位置的信號似乎依賴於間隙連接,因此在胚胎髮育的後期階段細胞的基本分化也是如此。還發現間隙連接是造成藥物產生影響的信號傳遞的原因,相反,一些藥物被證明可以阻斷間隙連接通道。
細胞死亡
“旁觀者效應”及其無辜旁觀者被殺的內涵也是由間隙連接介導的。當細胞由於疾病或損傷而受損並且開始死亡時,消息通過間隙連接被傳遞到連接到垂死細胞的相鄰細胞。這可能導致其他未受影響的健康旁觀者細胞也死亡。因此,在患病細胞中考慮旁觀者效應非常重要,這為更多資金和研究蓬勃發展開闢了道路。後來,對於受輻射或機械損傷損傷並因此傷口癒合的細胞,也研究了旁觀者效應。疾病似乎也會影響間隙連接在傷口癒合中發揮作用的能力。
雖然由於治療途徑的可能性而傾向於關注疾病中的旁觀者效應,但有證據表明在組織的正常發育中存在更重要的作用。一些細胞及其周圍基質的死亡可能是組織達到其最終構型所必需的,並且間隙連接對於該過程也是必不可少的。還有更複雜的研究試圖結合我們對間隙連接在傷口癒合和組織發育中的同時作用的理解。
間隙連接通過電和化學方式將細胞連接在大多數動物的身體上。電耦合可以相對快速地起作用。本節中的組織具有眾所周知的功能,觀察到由間隙連接協調,其中細胞間信號發生在微秒或更短的時間幀內。
心臟
縫隙連接在心肌中特別重要:收縮信號有效地通過間隙連接,使心肌細胞一致收縮。除了成人完全發育的骨骼肌和移動細胞類型(如精子或紅細胞)之外,間隙連接幾乎在身體的所有組織中表達。幾種人類遺傳疾病與間隙連接基因的突變有關。其中許多影響皮膚,因為這種組織嚴重依賴於間隙連接通訊來調節分化和增殖。心臟間隙連接可在藥理學上用rotigaptide打開。
神經元
位於神經元中的間隙連接通常被稱為電突觸。在描述間隙連接結構之前,使用電測量發現電突觸。電突觸存在於整個中樞神經系統中,並且已在新皮質,海馬,前庭神經核,丘腦網狀核,藍斑,下橄欖核,三叉神經中腦核,腹側被蓋區,嗅球,視網膜和脊椎動物的 脊髓。
對藍斑和神經 膠質細胞與Bergmann神經膠質細胞之間的小腦神經 膠質細胞偶聯有一些弱神經元的觀察。似乎星形膠質細胞通過間隙連接與其他星形膠質細胞和少突膠質 細胞偶聯。此外,間隙連接基因Cx43和Cx56.6中的突變導致白質變性,類似於在Pelizaeus-Merzbacher病和多發性硬化中觀察到的。
在神經元間隙連接處表達的連接蛋白包括:
1.m CX36
2.mCX57
3.m CX45
用mRNA檢測至少五種其他連接蛋白(m Cx26 ,m Cx30.2 ,m Cx32 ,m Cx43 ,m Cx47 )但沒有超微結構限定的間隙連接中相應蛋白的免疫細胞化學證據。那些mRNA似乎被細胞類型和細胞譜系特異性的微干擾RNA( miRNA )下調或破壞。
視網膜
視網膜內的神經元在一種細胞類型的群體內和不同的細胞類型之間顯示出廣泛的偶聯。
基本信息
- 中文名
- 間隙連接
- 外文名
- gap junction
- 含義
- 連接子進行的細胞間連接
- 類別
- 生物
- 性質
- 科學
- 組織層次
- dNA轉化為連接蛋白的RNA