心肌細胞
心肌細胞
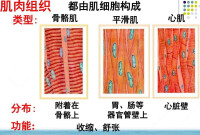
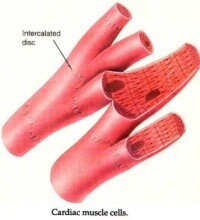
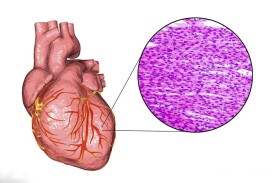
心肌細胞又稱心肌纖維,有橫紋,受植物性神經支配,屬於有橫紋的不隨意肌,具有興奮收縮的能力。呈短圓柱形,有分支,其細胞核位於細胞中央,一般只有一個。各心肌纖維分支的末端可相互連接構成肌纖維網。廣義的心肌細胞包括組成竇房結、房內束、房室交界部、房室束(即希斯束)和浦肯野纖維等的特殊分化了的心肌細胞,以及一般的心房肌和心室肌工作細胞。
心肌細胞
一類是普通的心肌細胞,包括心房肌和心室肌,含有豐富的肌原纖維,執行收縮功能,故又稱為工作細胞。工作細胞不能自動地產生節律性興奮,即不具有自動節律性;但它具有興奮性,可以在外來刺激作用下產生興奮;也具有傳導興奮的能力,但是,與相應的特殊傳導組織作比較,傳導性較低。
另一類是一些特殊分化了的心肌細胞,組成心臟的特殊傳導系統;其中主要包括P細胞和哺肯野細胞,它們除了具有興奮性和傳導性之外,還具有自動產生節律性興奮的能力,故稱為自律細胞,它們含肌原纖維甚小或完全缺乏,故收縮功能已基本喪失。還有一種細胞位於特殊傳導系統的結區,既不具有收縮功能,也沒有自律性,只保留了很低的傳導性,是傳導系統中的非自律細胞,特殊傳導系統是心臟內發生興奮和傳播興奮的組織,起著控制心臟節律性活動的作用。
心肌細胞
心肌細胞
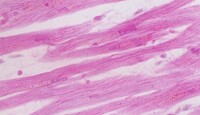
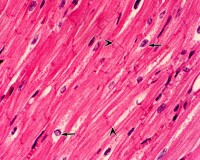
3.在電子顯微鏡下觀察,也可看到心肌細胞的肌原纖維、橫小管、肌質網、線粒體、糖原、脂肪等超微結構。但是心肌細胞與骨骼肌有所不同;心肌細胞的肌原纖維粗細差別很大,介於0.2~2.3微米之間;同時,粗的肌原纖維與細的肌原纖維可相互移行,相鄰者又彼此接近以致分界不清。心肌細胞的橫小管位於Z線水平,多種哺乳動物均有縱軸向伸出,管徑約0.2微米。而骨骼肌的橫小管位於A-I帶交界處,無縱軸向伸出,管徑較大,約0.4微米。心肌細胞的肌質網叢狀居中間,側終池不多,與橫小管不廣泛相貼。
心肌細胞
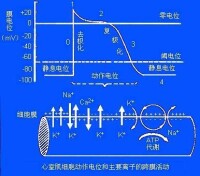
(一)心室肌細胞跨膜電位及其產生機理:
1.靜息電位:心室肌細胞在靜息時,細胞膜處於內正外負的極化狀態,其主要由K+外流形成。
2.動作電位:心室肌動作電位的全過程包括除極過程的0期和復極過程的1、2、3、4等四個時期。
0期:心室肌細胞興奮時,膜內電位由靜息狀態時的-90mV上升到+30mV左右,構成了動作電位的上升支,稱為除極過程(0期)。它主要由Na+內流形成。
1期:在復極初期,心室肌細胞內電位由+30mV迅速下降到0mV左右,主要由K+外流形成。
2期:1期復極到0mV左右,此時的膜電位下降非常緩慢它主要由Ca2+內流和K+外流共同形成。
3期:此期心室肌細胞膜復極速度加快,膜電位由0mV左右快速下降到-90mV,歷時約100~150ms。主要由K+的外向離子流(Ik1和Ik、Ik也稱Ix)形成。
4期:4期是3期復極完畢,膜電位基本上穩定於靜息電位水平,心肌細胞已處於靜息狀態,故又稱靜息期。Na+、Ca2+、K+的轉運主要與Na+--K+泵和Ca2+泵活動有關。關於Ca2+的主動轉運形式目前多數學者認為:Ca2+的逆濃度梯度的外運與Na+順濃度的內流相耦合進行的,形成Na+-Ca2+交換。(二)蒲肯野細胞的跨膜電位及產生機理:
蒲肯野細胞的動作電位及其產生機理與心室肌細胞基本相似,但其有4期自動除極化。4期自動除極化是膜對Na+通透性隨時間進行性增強(If內向電流)的結果。If通道與快Na+通道的主要區別是:①If的通道對離子的選擇性不強,雖然主要選擇的是Na+,但還有K+參與。而快Na+通道的選擇性強,主要允許Na+通透。②If的通道在復極達-60mV左右被激活,而快Na+通道在膜內電除極達-70mV左右被激活。③If的通道可被銫(Cs)所阻斷,而快Na+通道可被河豚毒阻斷。
(三)竇房結P細胞跨膜電位及產生機理:
1.P細胞動作電位的主要特徵4期膜電位不穩定,可發生自動除極,這是自律細胞動作電位最顯著的特點。
此外:
1)除極0期的峰值較小,除極速度較慢,約為10V/s,0期除極只到0mV左右。
2)復極由3期完成,基本沒有1期和2期。
3)復極3期完畢後進入4期,這時可達到的最大膜電位值,稱為最大舒張電位(或稱最大復極電位),約為-70mV。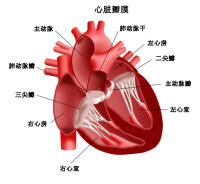
心肌細胞
2.P細胞動作電位的形成及離子流的活動
(1)0期除極的形成:0期除極的內向電流主要是由鈣離子負載的。
(2)3期復極的形成:0期除極后,慢鈣離子通道逐漸失活。3期是由鈣離子內流和鉀離子外流共同作用的結果。
(3)4期自動除極的形成:目前研究與三種離子流有關。
A:鉀離子外流的進行性衰減;
B:鈉離子內流的進行性增強;
C:生電性Na+--Ca2+離子交換。
(四)心肌細胞的電生理學分類
心肌細胞除了解剖生理特點分為工作細胞(非自律細胞)和自律細胞外,還可根據心肌細胞動作電位的電生理特徵(特別是0除極速率),把心肌細胞所產生的動作電位分為兩類:快反應電位和慢反應電位,而把具有這兩不同電位的細胞分別稱為快反應細胞和慢反應細胞:
1.快反應細胞包括:心房肌、心室肌和蒲肯野細胞,其動作電位特點是:除極快、波幅大、時程長。
2.慢反應細胞包括竇房結和房室交界區細胞,其動作電位特點是:除極慢、波幅小、時程短。
心肌細胞分類小節如下:
自律細胞
快反應自律細胞:如蒲肯野氏細胞
慢反應自律細胞:竇房結和房室交界區(房結區,結希區)細胞
非自律細胞快反應非自律細胞:心房肌、心室肌細胞
慢反應非自律細胞:結區細胞
美國科學家在《自然》(Nature)雜誌上發表研究報告指出,發現了一組可培植心肌細胞的幹細胞。帶領這項研究的科學家正是華人WilliamPu。美國麻省波士頓兒童醫院的研究人員表示,新發現的幹細胞位於心臟最外層的心外膜,或能修復已受損害的心臟組織。WilliamPu稱:“當病人心臟出現問題時,便會失去驅動心跳的心肌細胞。唯一的補救方法就是製造更多這類細胞。”據悉,研究人員是在偶然的情況下發現新幹細胞的。他們當時正在研究心外膜的另一組基因,所以要在活老鼠的胚胎上,用紅色熒光蛋白複合體標籤特定的細胞。出乎意料之外,他們竟然目睹心外膜細胞轉化成心肌細胞。WilliamPu的研究成果顯示,用基因編號為“Wt1”的幹細胞能製造出心肌細胞、滑肌細胞及內皮細胞
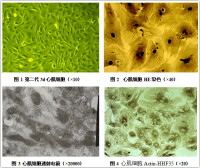
心肌細胞
一、心肌細胞肥大的概念,心肌組織包括心肌細胞和間質兩部分,其中心肌細胞占心臟總體積的75%;間質佔25%。心肌細胞肥大是指心肌細胞體積增大直徑增寬或長度增加和肌節數量增多,心肌過度肥大時可有心肌細胞增生。
二、心肌細胞肥大的細胞學基礎,心肌細胞是一種高度分化的終末細胞,其收縮蛋白以α-肌球蛋白(α-MHC)為主,主管收縮功能。收縮蛋白包括肌球蛋白,肌動蛋白、向肌球蛋白及向寧蛋白。其中肌球蛋白包括2個重鏈(MHC)、2個輕鏈(MLC),心臟僅有兩種MHC基因表達即α-MHC和β-MHC形成α-α、β-β同二聚體及α-β異二聚體,分別形成同功酶V1、V2及V3。正常情況下,胚胎心房及成人心房α-MHC即V1同功酶佔優勢,而左右心室從胚胎到成人β-MHC始終保持在80%~90%,以V3同功酶佔優勢。心肌細胞一般不能增殖,只有細胞體積的肥大,處於收縮狀態。胚胎期心肌細胞來源於肌幹細胞,經肌母細胞逐漸分化成成熟的心肌細胞,其收縮蛋白以β-MHC佔優勢,處於“合成狀態”,心肌肥大是心肌細胞從成熟的“收縮狀態”向“胚胎型合成狀態”轉化的一種現象。心肌細胞肥大時,其表型變化,體積增大,心肌細胞內收縮蛋白類型改變,同時也可有心肌間質細胞增殖。心肌細胞和間質細胞的生長有各自的調控機制,心肌細胞肥大可伴有也可不伴有間質細胞的增殖。
三、心肌細胞肥大的原因和機制 1958年Teare對肥厚性心肌病進行了描述,從此,人們一直對心肌肥厚的發生機制進行研究,近年研究表明心肌肥厚是一種複雜的多種因素參與調節的動態過程。心肌細胞肥大發生的生化基礎是心肌蛋白合成的增加,導致細胞體積的增加。各種機械刺激,化學因素作用都可導致心肌肥大。
1.機械性刺激的直接作用,長期的壓力和/或容積負荷過度。使心室壁應力增加,導致心肌肥大。整體實驗表明心臟受到負荷刺激時可發生心肌肥大。機械性刺激可通過促進蛋白質合成增加或/和促使蛋白質降解減少而導致心肌肥大。其機製為(1)細胞內CAMP增加在搏動或停搏的心臟,其主動脈壓從7.98kPa(60mmHg)升至15.96kPa(118mmHg)時,其蛋白質合成增加,核蛋白形成增多,CAMP含量增多及CAMP依賴性蛋白激酶活性增加,提示動脈壓力增高可通過CAMP依賴性作用機制加速蛋白質合成。(2)細胞內肌醇磷酯增加Portzer等報告,主動脈狹窄造成左室肥大時,肥大心肌的細胞漿蛋白激酶(PKC)活力較對照增加15%,細胞膜PKC活力增加40%。說明心肌牽張增加心肌內肌醇磷酯含量的原因可能是由於磷脂酶C的激活。(3)原癌基因表達增加 在壓力負荷過重所致心肌肥大早期,可觀察到原癌基因表達增加。(4)肌動蛋白和肌球蛋白基因表達增加 培養的心肌細胞持續受到牽張刺激時,其β-MHC和α-肌動蛋白的基因表達增加。(5)其它 心肌細胞間鈣離子通道,鈉離子內流及細胞內環境鹼性比等均可在牽張刺激所致的心肌肥厚中起重要調節作用。
心肌細胞

心肌細胞
在國外,心肌細胞電生理學的著作,日益增多,而在國內,則寥寥無幾,而為臨床醫生所讀的著作,更是缺如。
心肌細胞
方法:採用胰酶消化及新生大鼠心肌細胞與非心肌細胞如成纖維細胞、血細胞等的差時貼壁法分離提純心肌細胞,建立心肌培養模型。
結果:心肌細胞培養最長成活時間達3天,心肌細胞受消化時間的影響有不同的形態表現。
結論:可以利用差時貼壁法分離新生鼠心肌細胞並在體外進行培養,胰酶消化以低濃度短時間為佳。
1.材料和方法
1.1心肌細胞的消化
取一隻加蓋燒杯,內放一塊用乙醚飽和的紗布,把0~3日齡的大鼠幼鼠放進該燒杯中麻醉,用2%碘酒和75%酒精消毒胸腹皮膚,在無菌條件下開胸取出心臟,立即置於4ºCD-Hanks液(mmol/L:Nacl137,Kcl5.4,Na2HPO40.37,K2HPO40.44,NaHCO34.2)中剪取心室肌,洗凈殘血,剪成約1mm³大小的組織塊,棄去D-Hanks液加入0.08%胰蛋白酶液10~15ml,於37°C靜置5min,吸出上層懸液,並加入等量的含血清培養基。經終止消化后離心(1800r/min)棄上清液,加入含血清的培養液。吹散沉澱細胞,同條件下離心,用10%血清培養液製成細胞懸液,置於37°C含5%CO2培養箱中。
1.2心肌細胞分離
根據心肌細胞和非心肌細胞貼壁時間的不同採用2小時差速貼壁,分別獲得心肌成纖維細胞和心肌細胞。心肌細胞按1*106個細胞/ml總重50ml培養液中,培養的前2天在心肌細胞培養液中加入5-溴脫氧嘧啶核苷0.1mmol/l,以抑制非心肌細胞增殖,所有培養的心肌細胞均每2天換液一次。
1.3心肌細胞的無血清培養
當心肌細胞培養24小時后換無血清培養液(含DMEM培養液,胰島素10g/ml,鐵蛋白10g/ml,維生素C100g/L維生素B121.5µmol/L)繼續培養48小時,每隔8小時換液一次,盡量保持各添加成分濃度不變,最後收集細胞,進行測定。
2.實驗結果
本次實驗中新生大鼠心肌細胞培養最長存活時間為3天。實驗採取低濃度,短時間胰蛋白酶消化的方法。消化時間不同,所得到的細胞有不同的形態變化:消化3~5分鐘,高倍鏡下觀察心肌細胞大部分呈新月形,並可見細胞在培養液中呈頭尾相連的過程,呈動態變化。而消化5~10分鐘的心肌細胞捲曲呈圓形,密度低,活動能力差。細胞培養約8小時後向一處聚集,彼此鉤連呈現高密度區,這可能與細胞之間形成連接有關。由於某種不明原因,多次培養過程中心肌細胞中都出現不明微生物,致使細胞在3天後基本死亡。
3.實驗討論
有很多文獻報道新生大鼠心肌細胞的培養時間為10~12天,本次實驗的培養時間為3天,與之比較成活時間較短,但本實驗可以說明低濃度胰蛋白酶消化液(0.08%)比一般濃度(0.125%)的消化時間效果好,消化時間在3~5分鐘內可減少細胞死亡率。實驗過程中出現的細胞向一處聚集的現象和可見的連續的形態變化,可以說明體外培養心肌細胞重新連接成更大單位細胞團是可能的,它們可能通過形態的變化相互鉤連,連接成網。
日美聯合研究成功培育心肌細胞
2010年8月8日。日本和美國的研究人員6日在《細胞》雜誌網路版上發表論文說,他們通過在纖維原細胞中植入特定的3種基因,成功培育出心肌細胞。
研究人員發現,在小鼠胚胎的心臟中,有3種基因是生成心肌細胞必不可少的。通過向纖維原細胞中植入這3種基因,可以獲得驅動心跳的心肌細胞。
研究人員指出,與利用誘導多功能幹細胞(iPS細胞)培育心肌細胞相比,這種方法更加安全、簡捷。
該項研究負責人日本慶應大學助教家田真樹說:“今後將確認是否可以用同樣方法製造出人類心肌細胞。如果可行,心肌梗塞患者將無需接受開胸手術,而只需通過導入這些基因,讓那裡的纖維原細胞直接生成健康的心肌細胞。”
基本信息
- 中文名
- 心肌細胞
- 結構特徵
- 短柱狀
- 支配神經
- 植物性神經
- 類型
- 橫紋肌
- 別名
- 心肌纖維
- 組成
- 竇房結,房內束,房室交界部