血型系統
血型系統
血型系統(blood group system)是根據紅細胞膜上同種異型(或表型)抗原關係進行分類的組合。紅細胞抗原決定簇可引起同種異型免疫應答,也可引起異種免疫應答。在鑒定人的血型時,一般是用特異性的人抗血清進行凝集反應。每一個血型系統都是獨立遺傳的,控制一個血型系統的遺傳基因大多是在同一條染色體上。目前發現的人類血型系統有35個,但在臨床試驗中,ABO血型系統和Rh血型系統的臨床意義最為重要。

表1 不同血型紅細胞上的抗原和血清中的抗體
血液有不同類型是奧國病理學家、免疫學家卡爾.蘭德斯泰納(一八六八—一九四三)在1902年提出的。他曾從自己和五位同事身上取得血液樣本,合成三十個樣本,進行觀察研究。他發現有的樣本成功混合,有的卻發生凝結(黏在一起」。他於是領悟,每個樣本的情況並不完全相同。有兩人的樣本,紅血球上有一種稱為「Anti-gen」(抗原)的物質,他於是以「A」作標記;另外兩人的樣本,另有一種「抗原」,他依字母順序,以「B」作標記;只有一人的樣本,A抗原和B抗原都沒有,但血清中卻有兩種抗體,他自己的血液也是如此,他於是以「O」(表示無抗原)作標記。後來,他發現有一群人的血液,既有A抗原,也有B抗原,他便叫它做AB型。

表2 基因型與紅細胞的抗原關係
紅細胞血型是1900年由奧地利的K.蘭德施泰納發現的。他把每個人的紅細胞分別與別人的血清交叉混合后,發現有的血液之間發生凝集反應,有的則不發生。他認為凡是凝集者,紅細胞上有一種抗原,血清中有一種抗體。如抗原與抗體有相對應的特異關係,便發生凝集反應。如紅細胞上有A抗原,血清中有A抗體,便會發生凝集。如果紅細胞缺乏某一種抗原,或血清中缺乏與之對應的抗體,就不發生凝集。根據這個原理他發現了人的ABO血型。後來他又把不同人的紅細胞分別注射到家兔體內,在家兔血清中產生了3種免疫性抗體,分別叫做M抗體、N抗體及P抗體。用這3種抗體,又可確定紅細胞上3種新的抗原。這些新的抗原與ABO血型無關,是獨立遺傳的,是另外的血型系統。而且M、N與P也不是一個系統。控制不同血型系統的血型基因在不同的染色體上,即使在一個染色體上,兩個系統的基因位點也相距甚遠,不是連鎖關係,因此是獨立遺傳的。
細胞膜上附著有糖類,它們以寡糖鏈或多糖鏈的形式存在,其主要作用就是細胞與細胞之間的識別。一個細胞要識別另一個細胞,它們的糖類必須是同一類的,因此就有了血型。
美國科學家皮特·達達莫博士認為,人類的血型是由進化決定的。
我們的4種血型——O型、A型、B型和AB型——並不是在所有的人身上同時出現,而是由於不斷進化和人們在不同氣候地區定居下來后逐漸形成。在寒冷的年代,由於草原上可供吃用的東西匱乏,游牧部落不得不去適應新地形所能提供的新食物。由於新的飲食結構出現,人的消化系統和免疫系統也會隨之有所變化,緊接著血型也會有所變化。
O型血的歷史最為悠久。它大約出現於公元前6萬至4萬年之間,當時的尼安德特人吃的是簡單的飯食:野草、昆蟲和從樹上掉下來猛獸吃剩下的果實。而4萬年前出現了克魯馬儂人,他們以狩獵為生。在獵光了所有的大野獸后,他們從非洲向歐洲和亞洲轉移。
A型血出現在公元前2.5萬年至1.5萬年之間。當時,我們的以果實為生的祖先逐漸變成雜食。隨著時間的推移,農耕成為住在現今歐洲土地上的人們的主要生產方式,野禽野獸開始接受馴養,人的飲食結構隨之發生變化。就是現在,絕大多數A型血的人都居住在西歐和日本。
B型血出現在約公元前1.5萬年至新紀元之間。當時東非的一部分人被迫從熱帶稀樹榦草原遷徙到寒冷而貧瘠的喜馬拉雅山一帶。氣候的變化便成了催生B型血的主要因素。這種血型一開始出現在蒙古人種身上,隨著他們後來不斷向歐洲大陸遷徙,結果今天有很多東歐人都是這個血型。
人體的4種血型中最後出現的為AB型,它的出現還不到1000年的時間,是“攜帶”A型血的印歐語民族和“攜帶”B型血的蒙古人混雜在一起后的產物。AB血型的人繼承了耐病的能力,他們的免疫系統更能抵抗細菌,但他們易患惡性腫瘤。
很快會出現第5種血型。完全有可能出現一種新血型,比如說C型。只有這種有新血型的人才能在人口過於稠密、自然資源所剩無幾的嚴重污染世界上生存下來,因為這時原先那4種血型,也就是說,有好幾十億甚至上百億的人將抵擋不住這種日益加劇的生態災難,他們會很快消失。這就是皮特,達達莫博士得出的結論。
血型系統
| 中文名稱 | 英文通用名稱 | 簡稱 | 編碼基因名稱 |
| ABO型血型系統 | ABO blood group system | ABO | ABO |
| MNS血型系統 | MNS antigen system | MNS | GYPA GYPB GYPE |
| P血型系統 | P antigen system | P1 | P1 |
| Rh血型系統 | Rhesus blood group system | RH | RHD RHCE |
| Lutheran血型系統 | Lutheran antigen system | LU | LU |
| Kell血型系統 | Kell antigen system | KEL | KEL |
| Lewis血型系統 | Lewis antigen system | LE | FUT3 |
| Duffy血型系統 | Duffy antigen system | FY | DARC |
| Kidd血型系統 | Kidd antigen system | JK | SLC14A1 |
| Diego血型系統 | Diego antigen system | DI | SLC4A1 |
| Yt血型系統 | Yt antigen system | YT | ACHE |
| XG血型系統 | XG antigen system | XG | XG MIC2 |
| Scianna血型系統 | Scianna antigen system | SC | ERMAP |
| Dombrock血型系統 | Dombrock antigen system | DO | ART4 |
| Colton血型系統 | Colton antigen system | CO | AQP1 |
| Landsteiner-Wiener血型系統 | Landsteiner-Wiener antigen system | LW | ICAM4 |
| Chido/Rodgers血型系統 | Chido/Rodgers antigen system | CH/RG | C4A C4B |
| Hh/孟買血型系統 | Hh/Bombay antigen system | H | FUT1 |
| Kx蛋白 | Kx protein | XK | XK |
| Gerbich血型系統 | Gerbich antigen system | GE | GYPC |
| Cromer血型系統 | Cromer antigen system | CROM | CD55 |
| Knops血型系統 | Knops antigen system | KN | CR1 |
| Indian血型系統 | Indian antigen system | IN | CD44 |
| Ok血型系統 | Ok antigen system | OK | BSG |
| Raph血型系統 | Raph antigen system | RAPH | CD151 |
| JMH血型系統 | John Milton Hagen antigen system | JMH | SEMA7A |
| Ii血型系統 | Ii antigen system | I | GCNT2 |
| Globoside血型系統 | Globoside antigen system | GLOB | B3GALT3 |
| GIL血型系統 | GIL antigen system | GIL | AQP3 |
簡介
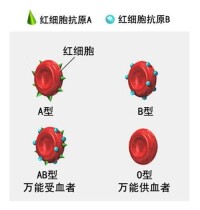
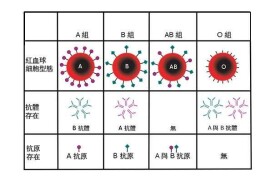
卡爾·蘭德施泰納(Karl Landsteiner)(1900年)發現的人類第一個血型系統。其紅細胞上的抗原和血清中的抗體如表1所示。
ABO血型由A、B和O三個等位基因控制遺傳。其中A和B基因是顯性基因,O基因是隱性基因。染色體上基因內容的組合(基因型或遺傳型)與紅細胞上表現的抗原(表型)關係如表2所示。
血型系統
ABO血型系統與其他血型系統不同,具有“天然”的抗體,例如,A型者有抗B抗體,B型者有抗A抗體,O型者有抗A抗B抗體。這種抗體是因出生後接受自然環境中具有同樣特異性的抗原性物質的隱性免疫作用而產生的。
血型的調查和建檔對於了解種族關係用處很大。
基因
科學研究發現,控制人類的ABO血型的遺傳基因有3個:IA、IB、i。
其中,IA和IB對i為顯性,IA、IB間無顯隱性關係。
也就是說:
A型血的基因組成可以是IAIA或IAi;
B型血的基因組成可以是IBIB或IBi;
AB型血的基因組成是IAIB;
O型血的基因組成是ii。
鑒定
通常只用兩種抗血清即抗A及抗B血清,就可將群體分為四種血型。
根據血型的遺傳規律,和臨床工作方例起見,配偶間所生子女的血型如下:
各種ABO配偶所生子女的血型:
| 婚配式 | 子女可能有的血型 | 子女不可能有的血型 |
| A*A | A、O | B、AB |
| A*B | A、B、AB、O | 無 |
| A*AB | A、B、AB | O |
| A*O | A、O | B、AB |
| B*B | B、O | A、AB |
| B*AB | A、B、AB | O |
| B*O | B、O | A、AB |
| AB*AB | A、B、AB | O |
| AB*O | A、B | AB、O |
| O*O | O | A、B、AB |
各種ABO配偶所生子女血型以及概率:
| 婚配式 | A型血的可能 | B型血的可能 | O型血的可能 | AB型血的可能 |
| A*A | 15/16 | 1/16 | ||
| A*B | 3/16 | 3/16 | 1/16 | 9/16 |
| A*O | 3/4 | 1/4 | ||
| A*AB | 1/2 | 1/8 | 3/8 | |
| B*B | 15/16 | 1/16 | ||
| B*O | 3/4 | 1/4 | ||
| B*AB | 1/8 | 1/2 | 3/8 | |
| O*O | 1 | |||
| O*AB | 1/2 | 1/2 | ||
| AB*AB | 1/4 | 1/4 | 1/2 |
輸血時凝集情況(第一行為供體,第一列為受體):
| 受體\供體 | A型 | B型 | AB型 | O型 |
| A型 | 不凝集 | 凝集 | 凝集 | 不凝集 |
| B型 | 凝集 | 不凝集 | 凝集 | 不凝集 |
| AB型 | 不凝集 | 不凝集 | 不凝集 | 不凝集 |
| O型 | 凝集 | 凝集 | 凝集 | 不凝集 |
表3&amp
RH陰性者不能接受RH陽性者血液,因為RH陽性血液中的抗原將刺激RH陰性人體產生RH抗體。如果再次輸入RH陽性血液,即可導致溶血性輸血反應。但是,RH陽性者可以接受RH陰性者的血液。
臨床意義
1.防止Rh血型系統所致的溶血性輸血反應:Rh陰性患者如輸入Rh陽性血液后便可刺激機體產生抗Rh抗體,當再次輸入Rh陽性血液時,就會發生溶血性輸血反應。如Rh陰性婦女曾孕育過Rh陽性胎兒,當輸入Rh陽性血時亦可發生溶血反應。所以需要輸血的患者和供血者,除檢查ABO血型外,還應做Rh血型鑒定,以避免這種情況的發生。
2.Rh陽性紅細胞引起的新生兒溶血症:Rh陰性的母親孕育了Rh陽性的胎兒后,胎兒的紅細胞若有一定數量進入母體時,即可刺激母體產生抗Rh陽性抗體,如母親再次懷孕生第二胎時,此種抗體便可通過胎盤,溶解破壞胎兒的紅細胞造成新生兒溶血。若孕婦原曾輸過Rh陽性血液,則第一胎即可發生新生兒溶血。
RH血型系統,其中含有5種抗原,即C、c、D、E、e(不存在d抗原)。其抗原強弱順序為D、E、c、C、e。凡紅細胞含D抗原者為Rh陽性,否則為陰性。Rh血型無天然抗體,其抗體多由輸血(Rh陰性者被輸入Rh陽性血液)或妊娠(Rh陰性母親孕育著Rh陽性胎兒)免疫生成,具有重要臨床意義。一旦形成抗體,如再輸入Rh陽性血液,可發生嚴重輸血反應。再孕育Rh陽性胎兒可發生新生兒溶血症。
因此RH陰性的女性在輸了RH陽型的血后,血液里產生了抗體,就不能再懷RH陽性的孩子了,否則嬰兒多半難以存活。也有部分存活胎兒由於溶血所產生的大量膽紅素進入腦細胞,引起新生兒中樞神經細胞病變,(稱為核黃疸。核黃疸殘廢率極高)即使倖存也會影響病兒的智力發育和運動能力。
女性如果不輸RH陽性的血,則可生育第一胎,這是由於第一胎懷孕時,孕婦體內產生的抗體量較少,還不足以引起胎兒發病。如果第一胎是RH陽性,那麼以後就不能繼續生育了。
如果男性是RH陰性,那麼生完RH陽性的孩子后也不要生育第二胎。但是男性輸完RH陽性的血后不會喪失生育能力。
RH陰性者自我保護
血液只能同型輸注,即A型Rh(-)患者只能輸A型Rh(-)血,B型的只能輸B型Rh(-)血,假如您生病或手術需要輸血時,您一定要將您是Rh(-)血型的情況告知醫生,以便醫生及早和市血站聯繫,組織您所需要的Rh(-)血源。假如您是未婚女性,請您做好計劃生育以避免人工流產,若您已有流產或輸血史,妊娠期務必到血站血型室進行新生兒溶血病的預測檢查,以防止今後新生兒溶血病的發生。
您知道血型工作者已經為您做好了哪些工作嗎?
緊急情況時能否及時向Rh(-)患者供應同型血液是一件生命攸關的大事,血站的血型工作者以保障每一位Rh(-)患者的輸血安全為己任,常年不懈地對數以萬計的供血人群進行Rh(-)血型篩選,一旦篩選到Rh(-)血,這袋血就立即做上特殊的標記並保存在4℃冰箱中以備急需。
RH陰性者輸血保障
儘管輸血工作者防患於未然已竭盡全力,但緊急情況時Rh(-)者可能缺乏同型血而危機生命的潛在危險依然存在。其實,消除這一困擾的方法很簡單:即所有Rh(-)者同舟共濟,在隨時作為可能的受血者的同時,也隨時作為一名高尚的供血者,這樣您和所有的Rh(-)者的生命都將得保障。有些血型抗體是不完全抗體,與相應的抗原細胞結合后看不出凝集現象,血清中有抗體但不容易發現。1945年抗人球蛋白試驗應用到血型檢查中來,這種試驗就可檢查不完全抗體,從此,許多血型抗原陸續被人發現。每當發現一個新抗原后就要確定這一抗原與已經發現的血型是什麼關係,這樣在人的紅細胞上便確定了若干血型系統。此外,還有一些抗原,或因其在群體中出現的頻率太高,或因其在群體中分佈的頻率太低,對它們無法進行遺傳學分析。在沒有弄清它們的遺傳關係以前,暫且把這些抗原分別叫做高頻率抗原及低頻率抗原,對於它們的歸屬有待進一步確定。
紅細胞膜中夾雜著3種蛋白質:糖蛋白、簡單蛋白及膜收縮蛋白。紅細胞抗原有些突出在細胞表面,好像伸出在地面上的樹枝,如ABH抗原;有些鑲嵌在細胞膜內,如Rh抗原。抗原與抗體發生特異反應的部分,叫做抗原決定簇。血型抗原決定簇的化學組成,有的已經清楚,但大部分不清楚。有些血型在體液中存在可溶性抗原,叫做血型物質。從人體分離出來的ABH及Lewis血型物質是糖蛋白,即在肽鏈的骨架上連接著一些糖的側鏈,這些糖鏈便是特異性決定簇。ABH及Lewis血型物質的特異性決定簇很相似,只是在糖鏈上個別糖的種類或同一種糖由於存在位置不同,就顯出不同的特異性。比如A與B的抗原特異性,只是在糖鏈上有一個糖不相同,便顯示出不同的特異性。A抗原決定簇在糖鏈的終末端是一個N-乙醯半乳糖胺,而B抗原決定簇在糖鏈的終末端卻是一個D-半乳糖。
紅細胞上的ABH抗原決定簇,雖與體液中的抗原決定簇糖鏈結構相同,但連接的骨架不同。紅細胞上的糖鏈是通過神經鞘氨醇與脂肪酸結合在一起,而不是與蛋白質結合在一起,所以紅細胞上的ABH抗原是糖脂而不是糖蛋白。
MN·P及I血型的抗原決定簇也是碳水化合物。Rh抗原的決定簇可能是蛋白質,因為紅細胞經硫氫化物、脲素及蛋白酶等物處理后,Rh活性即行消失。
有一些血型抗體,如抗IH,抗IA,抗IB,抗IP1等,只與帶有I抗原及另外一個抗原的細胞發生反應,而不與其中只有一個抗原的細胞發生反應。說明這些抗原為複合抗原,在一個分子上具有兩種特異性。
Lewis血型抗原實際上是血漿中的抗原,紅細胞上的Lewis抗原是從血漿中吸附來的。I抗原在分泌液中雖有可溶性抗原,但不存在於血漿中。另外有些血型是在血漿中存在可溶性抗原,分泌液中卻不存在。Bg抗原實際是白細胞的抗原,可能從白細胞脫落到血漿中,再從血漿中吸附到紅細胞上,表現為紅細胞的抗原。Chido血型及Rodger血型的抗原與血漿中的補體第四成分(C4)有關。用電泳方法分析人的C4,可以見到3種類型:泳動快的(F);泳動慢的(S);快慢兩種成份都有的(FS)。血漿中只有F成份的人,紅細胞上有Rodger抗原。只有S成份的人,紅細胞上有Chido抗原。兩種成份全有的人,紅細胞上也同時具有Chido及Rodger兩種抗原。
各種血型抗原在紅細胞上的分佈是不同的,有的密集,有的疏鬆。抗原數目的多少決定了抗原的強弱。用放射性碘標記的兔抗A及抗B血清,檢查人的紅細胞,根據每個細胞上的放射性強度,可推算出每個紅細胞上的抗原數目。
各種血型抗原在個體發育不同階段強度是不相同的。新生兒的ABO及Lewis抗原與其相應的抗體之反應較成人細脆弱。不到10厘米的胎兒之紅細胞就能與抗P1血清發生反應,但其反應強度較成人紅細胞弱。新生兒的紅細胞吸收抗I的能力幾乎與成人紅細胞一樣,但凝集反應強度遠較成人紅細胞弱。可是與抗i血清的凝集卻比成人紅細胞強。Yta及Xga抗原在新生兒紅細胞上稍較成人紅細胞弱,而Rh、Kell、Duffy、Jk、MNSs、Di及Do等系統的抗原在出生時已發育完全。Chido血型的抗原在新生兒血漿中可以檢出,但在紅細胞上不能發現。
抗體是免疫球蛋白,但不一定所有免疫球蛋白都是抗體。只要具有抗體結構的糖蛋白便為免疫球蛋白。免疫球蛋白以Ig表示,現已發現人類具有五類免疫球蛋白,分別叫做IgG、IgM、IgA、IgD及IgE。與血型有關的免疫球蛋白只有三類,即IgG、IgM及IgA三類。
根據抗體在體內出現是否有可查覺的抗原刺激,有所謂“天然抗體”及“免疫性抗體”之分。凡未經抗原刺激就在體內的血清中出現的抗體,叫做“天然抗體”;機體受同種或異種抗原的刺激后血清中所產生的抗體,叫作免疫性抗體。
對於“天然抗體”的產生有兩種解釋:一種說法認為在體內存在“抗原致敏”細胞,不需要抗原刺激就能產生特異性抗體;另一種解釋認為“天然抗體”是異種凝集素,周圍環境存在著一些與血型抗原相似的物質,機體接觸這些物質后,所產生的交叉反應抗體。比如某些細菌含有與人的A,B抗原相似的抗原,當人們吸入或吞下這些細菌后,便產生交叉反應抗體。
“天然抗體”在低溫與其相應的抗原細胞反應強,有很多“天然抗體”當溫度超過25℃時即無活性。有的“天然抗體”有結合補體的能力,有的則沒有。如Lewis血型抗體幾乎都有結合補體的能力,而抗M及抗N就沒有結合補體的能力。“天然抗體”通常是IgM免疫球蛋白,但有些“天然抗體”卻是IgG免疫球蛋白。如有的抗Lea,抗M、抗N及抗K“天然抗體”是IgG。
免疫性抗體則是指機體受同種或異種抗原刺激后處於超免疫狀態而產生的抗體。輸血、妊娠是產生同種免疫抗體的主要原因。接受菌苗、抗血清(白喉、破傷風抗毒素)注射,以及使用過豬的胃、肝浸液的人,血清中的抗A、抗B效價升高,是異種免疫引起的免疫性抗體的例子。免疫性抗A及抗B,在許多方面與“天然抗體”不同。
有的抗體與其相應的抗原細胞在鹽水介質中即可出現凝集,這樣的抗體稱為完全抗體;有的抗體在鹽水介質中只能與其相應的抗原細胞結合(致敏),但不能出現凝集,這樣的抗體稱為不完全抗體。欲使不完全抗體與其相應的抗原細胞出現凝集,還需藉助其他介質,如酶處理紅細胞,或將紅細胞懸浮在大分子膠體液中,或利用抗球蛋白血清的幫助。實際上完全抗體一般是指IgM類型的抗體,而不完全抗體多為IgG類別的抗體。IgA主要在分泌液中,在血型抗體中不佔主要位置。
ABO血型
物質除存在於紅細胞膜上外,還出現於唾液、胃液、精液等分泌液中。中國60%漢族人唾液中有ABO血型物質。血型物質的化學本質是指構成血型抗原的糖蛋白或糖脂,而血型的特異性主要取決於血型抗原糖鏈的組成(即血型抗原的決定簇在糖鏈上)。A、B、H3種血型抗原化學結構的差異,僅在於糖鏈末端的1個單糖。A抗原糖鏈末端為N-乙醯半乳糖,而B抗原糖鏈末端為半乳糖,H抗原和A、B抗原相比則糖鏈末端少1個半乳糖或N-乙醯半乳糖。1981年已有人用綠咖啡豆酶(半乳糖苷酶)作用於B型紅細胞,切去B抗原上的半乳糖,從而使B型轉變成O型獲得成功。
E.von鄧格恩及L.希爾斯費爾德於1911年發現A血型的亞型。他們看到不同A型人的紅細胞與抗A血清發生凝集反應的強度不一,在反應弱的A型人血清中還有一種抗體能與反應強的A型紅細胞發生凝集反應。據此認為在A型中存在亞型;即A1及A2亞型。A1.型紅細胞與抗A血清(來自B或O型人)反應強,而A2型紅細胞與抗A血清反應弱。而且在部分A2型人的血清中,除存在的抗B外,還有不規則的抗A1。在B型人血清中有兩種抗體:抗A及抗A1。抗A能與A1及A2細胞發生反應;抗A1隻與A1細胞發生反應。A1型紅細胞上有A及A1兩種抗原。A2細胞上只有A抗原。AB型也可分為A1B及A2B等亞型。此外還有一些其他亞型。
MN血型
紅細胞膜上另一類血型抗原叫MN抗原,即紅細胞膜上的血型糖蛋白A。它在SOS凝膠電泳譜上顯示兩條區帶,即PAS-1和PAS-2,血型糖蛋白A是兩者的二聚物。已知血型糖蛋白A由131個氨基酸組成,其一級結構已測定(圖2)。血型糖蛋白A的肽鏈呈三節式結構,中間第73~92號氨基酸為疏水性肽鏈,可橫穿膜脂層;N端肽鏈位於膜外側,與血型活性有關,在這段肽鏈上分佈有15條O-糖苷鍵型糖鏈和1條N-糖苷鍵型糖鏈,糖鏈中唾液酸占紅細胞膜上全部唾液酸的一半以上;C端肽鏈位於膜內側,含較多酸性氨基酸。
MN抗原由M抗原和N抗原兩部分組成,如果用神經氨酸酶將M抗原切去1個唾液酸(N-乙醯神經氨酸),則為N抗原,如再切去一個唾液酸則抗原性完全失去。MN抗原的抗原性還和肽鏈上的氨基有關,若將氨基用乙醯基保護后即失去抗原性。
白細胞血型HLA
HLA是人類白細胞抗原中最重要的一類。與紅細胞血型相比,人們對白細胞抗原的了解較晚,人體第一個白細胞抗原Mac是1958年法國科學家J.多塞發現的。HLA是人體白細胞抗原的英文縮寫,已發現HLA抗原有144種以上,這些抗原分為A、B、C、D、DR、DQ和DP7個系列,而且HLA在其他細胞表面上也存在。
HLA抗原是一種糖蛋白(含糖為9%),其分子結構與免疫球蛋白極相似(圖3)。HLA分子由4條肽鏈組成(含2條輕鏈和2條重鏈),重鏈上連接2條糖鏈。HLA分子部分鑲嵌在細胞膜的雙脂層中,其插入膜的部分相當於免疫球蛋白IgG的Fc區段,輕鏈為β-微球蛋白。由於分子結構上的相似,故HLA與有保衛功能的免疫防禦系統密切相關。
此外,HLA和紅細胞血型一樣都受遺傳規律的控制。決定HLA型的基因在第6對染色體上。每個人分別可從父母獲得一套染色體,所以一個人可以同時查出A、B、C、D和DR5個系列中的5~10種白細胞型,因此表現出來的各種白細胞型有上億種之多。在無血緣關係的人間找出HLA相同的兩個是很困難的。但同胞兄弟姊妹之間總是有1/4機會HLA完全相同或完全不同。因此法醫鑒定親緣關係時,HLA測定是最有力的工具。
血型不僅在輸血上有重要意義,而且在人種學、遺傳學、法醫學、移植免疫、疾病抵抗力(或易感性)等方面都有應用價值,在輸血前,一定要檢查病人(受血者)和輸血人(供血者)的血型,並且要進行交叉配血試驗。在臨床醫學中,除輸血、移植免疫外,對新生兒溶血病、自身免疫性溶血性貧血特異性抗體的檢查,也都需要血型知識和有關技術。
稀有血型就是一種少見或罕見的血型。這種血型不僅在ABO血型系統中存在,而且在稀有血型系統中也還存在一些更為罕見的血型。隨著血型血清學的深入研究,科學家們已將所發現的稀有血型,分別建立起的稀有血型系統,如RH、MNSSU、P、KELL、KIDD、LUTHERAN、DEIGO、LEWIS、DUFFY以及其他一系列稀有血型系統。
還有一種叫孟買型的稀有血型系統,在這種血型的紅細胞上,沒有A、B和H抗原,但在血清中卻同時存在A、B和抗H三種抗體。
在稀有血型系統中,除RH血型系統外,其他各血型人數在總人口中所佔比例非常小。因此,它們在實際的臨床上遠沒有ABO及RH血型系統重要,但是,就其具體來說,如用血不當,有些抗體仍可出現致命的惡果。隨著社會的進步,人民生活水平的提高,開展稀有血型的檢測,建立完整的稀有血型檔案,對於保障廣大群眾的身體健康和適應我國改革開放形勢的需要,都具有深遠的意義。
基本上,O型是世界上最常見的血型。但在某些地方,如挪威,A型血型的人較多。A型抗原一般比B型抗原較常見。AB 型血型因為要同時有A及B抗原,故此亦是ABO血型中最少的。ABO血型分佈跟地區及種族有關。
血型的性格在幼兒期、少年期、青春期、中年期、老年期各有不同。一個人走向社會,從參加工作后,由新手成為骨幹,由下級成為上級,由工作直到退休,這期間其性格也都在不斷地變化。在這種變化中,可以看到不同血型的許多特徵。當然,血型與性格之間也並不是必然聯繫,只能作為一種參考。
A型人小時候比較任性,年輕時性格果斷剛毅,時時處處要強。走向社會後,隨著年齡的增長和社會經驗的積累,他們開始克制自己的情緒,表現出穩重謙虛的態度,容易成為不願過份表現自己的謹慎派。A型人在老年時,則顯得很固執。
B型人大都有一個天真浪漫的幼年期,隨著年齡的增長,逐漸分成心直口快和不擅交際應酬型兩種傾向。B型人由於性格自幼到老變化不大,相對來說會讓人感到他們越活越年輕。
O型人年少時比較溫順,但隨著年齡的增長,他們會積極地呈現出強烈的自我主張和自我表現,甚至成為非常有魄力的人。O型人從小至老的變化是最大的,往往是少年溫順,老來強硬。
AB型人大多小時候怕陌生人,很閉塞,但長大以後善交朋友,交際廣泛。AB型人因過於自信,容易自滿,老年時給人感覺很傲慢。
過去人們認為只有人才有血型,已知狗、雞和許多動物都有血型系統。生長在美國緬因海灣的角鯊有4種血型。大馬哈魚至少有8種抗原類型或類型的組合。這些不同類型的出現通常隨不同地區的種群而異。家畜也有血型,馬有4種,牛有3種,豬也有4種。在人類學上,根據A型、B型及AB型三型的出現率的多少組成一個指數叫做種族生化指數來研究各種血型在各人種中的分佈規律。O型的高頻率分佈在歐洲西北部、西南非、部分澳大利亞及南印度和中美洲;B型的最高頻率分佈於中亞及北印度;A型在歐洲、西亞及澳大利亞南部的土著中的是最高的,而在某些美洲印第安人部族中是最高的。
靈長類的血型可以通過抗A和抗B血清來測定。黑猩猩的血全部屬於O型或A型,猩猩屬於B型,大猩猩有B型也有A型,長臂猿血型有A型、B型及AB型。低等靈長類在紅血球里沒有抗原,但在它們的唾液里分泌ABO抗原。舊大陸猴大多數是血型A型,新大陸猴血型也是A型,但個別的在唾液里有象B一樣的抗原。在某些靈長類中發現具有類似人類M的抗原,如在黑猩猩體內發現了具有M血型和N血型,在靈長類中也發現具Rh抗原的。
基本信息
- 中文名
- 血型系統
- 外文名
- blood group system
- 定義
- 根據紅細胞膜上同種異型抗原關係進行分類的組合
- 類型
- 遺傳性狀
- 形式
- 血液抗原
- 根據
- 紅細胞膜上同種異型抗原關係
- 用途
- 鑒定人的血型