光敏素
植物體內的蛋白
光敏素
不論是黃色或綠色組織中都至少存在兩種類型的光敏色素分子:光敏色素Ⅰ(PⅠ)和光敏色素(PⅡ)。前者為光不穩定型、後者為光穩定型。nPⅠ在黃化組織中大量存在,在光轉變成Pfr后就迅速降解,因此在綠色組織中含量較低。PⅡ在黃化組織中含量較低,僅為PⅠ的1%~2%,但光轉變成Pfr后較穩定,加之在綠色植物中PⅠ被選擇性降解,因而PⅡ雖然含量低,卻是綠色植物中主要的光敏色素。兩類光敏色素分別調控不同的生理反應。一般認為PⅠ參與調控的反應時間較短,而PⅡ參與調控的反應時間較長。PⅡ調控的反應有時不被遠紅光和暗期所逆轉
光敏素分子由蛋白質和生色團兩部分組成。單體(1分子色素和1分子蛋白)的分子量在不同植物中有些不同,變化在 50~150之間。例如燕麥的為60,黑麥的為 120。光敏素的蛋白質中有很大比例的酸性和鹼性氨基酸,以及含硫氨基酸,如半胱氨酸。所以光敏素是一個高度帶電、非常活潑的蛋白分子,能通過內部重組而改變形式。它的等電點的pH值為6。
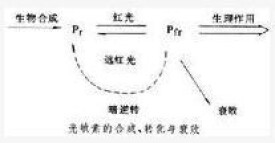
光敏素的生色團的每個亞基含有一個開鏈的四吡咯輔基。生色團以共價鍵與蛋白質相連。光敏素的兩種不同形式(Pr與fr)的吸收光譜不同。Pr(r=red,紅)型為藍色,吸收峰在波長660納米的紅光部分;fr(fr=farred,遠紅)型為黃綠色,吸收高峰在波長730納米的遠紅光部分。兩種形式的光敏素吸收相應波長的光後分別轉化成另一種形式。由於兩種形式的吸收光譜有部分重疊,只是吸收率不同,所以在不同波長的光下進行不同程度的相互轉化,形成不同的Pr/fr比值。因Pr對730納米或更長遠紅光吸收極少,故在遠紅光下fr向Pr的轉化接近完全;而Pr的吸收峰660納米處fr卻也有少量吸收,所以在紅光下最多只能將80%左右的Pr轉化為fr。
在調節植物光形態發生中起作用的是Pfr,Pr則為生理鈍化型。光敏素在調節種子萌發、莖葉生長、下胚軸“鉤”的伸直、葉綠體運動、花芽分化以及塊莖塊根形成中起作用。在種子萌發期和幼苗發育期以及營養生長向生殖生長轉化的時期,光敏素的調節作用最為明顯。
紅光促進萵苣種子等需光種子的萌發。有些種子植物幼苗出土見光后,下胚軸停止生長,下胚軸“鉤”伸直,子葉展開。在日光下生長的植物,莖的伸長受抑制;暗期光間斷抑制短日植物開花,而促進長日植物開花,這些現象中起作用的都是光下特別是紅光下形成的fr。
除了光形態發生效應以外,光敏素還參與調節膜的性質、50多種酶的活動、葉綠素和花青素的合成、乙醯膽鹼的含量、合歡等豆科植物的小葉的開合等。此外,光敏素還明顯激活rRNA順反子的轉錄,加強rRNA前體的形成,以及控制聚核糖體的形成和核糖體的功能。
光敏素在體內合成時,Pr形式先形成,在光下一部分轉變為fr。在黑暗中生長的植物,如黃化幼苗,只含Pr。fr除在遠紅光下向 Pr轉化外,還會“衰敗",失去活性。此外,在有些植物中,fr在暗中也會轉化為Pr,這個過程稱為光逆轉(見圖)。在Pr和fr的相互轉化中,還有一系列中間狀態,其中有的具有生理活性。在連續光照下,光敏素的各種形式之間達到恆定的比例,Pr合成速度等於fr的衰敗速度,光敏素的總量則恆定。離體的光敏素的兩種形式也可以在不同波長的光下相互轉化,但所達到的Pr/fr比值與同樣光下體內的比值不同,說明光敏素在體內的存在狀態與離體時不完全相同。由於光敏素的形式和狀態的多樣性和其間轉化速率受各種條件影響的複雜性,使得光敏素在不同植物、不同器官中對各種生長發育過程的調節有許多複雜的情況。光敏素還與植物體內生物鐘協同作用,賦予植物以精確計量日照長度的本領。
基本信息
- 外文名
- phytochrome