聚核糖體
聚核糖體
[編輯本段]核糖體(ribosome)定義
核糖體是細胞內一種核糖核蛋白顆粒(ribonucleoproteinparticle),主要由rRNA和蛋白質構成,其惟一功能是按照mRNA的指令將氨基酸合成蛋白質多肽鏈,所以核糖體是細胞內蛋白質合成的分子機器。
[編輯本段]核糖體蛋白
構成核糖體的蛋白質。大腸桿菌核糖體蛋白的初級結構均被確定。大腸桿菌核糖體的30S亞基含S1—S21共21種蛋白質,50S亞基含L1—L34共34種蛋白質。這些蛋白質已被全部分離純化。分子量約1萬到3萬。除S6、L7、L12之外全是鹼性蛋白質。這些蛋白質是免疫學上獨立的蛋白質,只有L7、L12顯示出相互交叉反應。已知L7與L12是同一蛋白質,L7的N末端被乙醯化。已經確定了幾種蛋白的一級結構。機能已經明確的蛋白質如下述:S1:與蛋白質合成的i因子(干擾因子)和Qβ複製酶的亞基Ⅰ為同一物質,可與mRNA結合;S4:ram(核糖體的雙關性ribosomalambiguity)基因的產物;S5:SPc〔壯觀黴素(Spectinomycin)抗性〕基因的產物;S12:str(鏈黴素抗性)基因的產物;L7、L12:有和多肽鏈延長因子Tu及G間的相互作用,也有和起始因子和終止因子的相互作用。L11:肽基轉移酶。
[編輯本段]核糖體的形成
真核細胞的大小亞基是在核中形成的,在核仁部位rDNA轉錄出45SrRNA,是rRNA的前體分子,與胞質運來的蛋白質結合,再進行加工,經酶裂解成28S,18S和5.8S的rRNA,而5SrRNA則在核仁外合成28S,5.8S及5SrRNA與蛋白質結合,形成RNP分子團。為大亞基前體,分散在核仁顆粒區,再加工成熟后,經核孔入胞質為大亞基,18SrRNA也與蛋白質結合,經核孔入胞質為小亞基。大小亞基在胞質中可解離存在,在需要時也可在>0.001MMg存在時,但合成完整單核糖體,才具有合成功能,當MG4<0.001M時則又重新解離。
[編輯本段]構成核糖體的蛋白質
按核糖體存在的部位可分為三種類型:細胞質核糖體、線粒體核糖體、葉綠體核糖體。
按存在的生物類型可分為兩種類型:真核生物核糖體和原核生物核糖體。
原核細胞的核糖體較小,沉降係數為70S,相對分子質量為2.5x103kDa,由50S和30S兩個亞基組成;而真核細胞的核糖體體積較大,沉降係數是80S,相對分子質量為3.9~4.5x103kDa,由60S和40S兩個亞基組成。典型的原核生物大腸桿菌核糖體是由50S大亞基和30S小亞基組成的。在完整的核糖體中,rRNA約佔2/3,蛋白質約為1/3。50S大亞基含有34種不同的蛋白質和兩種RNA分子,相對分子質量大的rRNA的沉降係數為23S,相對分子質量小的rRNA為5S。30S小亞基含有21種蛋白質和一個16S的rRNA分子。
真核細胞中,核糖體進行蛋白質合成時,既可以遊離在細胞質中,稱為遊離核糖體(freeribosome)。也可以附著在內質網的表面,稱為膜旁核糖體或附著核糖體。參與構成rER,稱為固著核糖體或膜旁核糖體,是以大亞基圓錐形部與膜接著遊離核糖體(freeribosome)。。分佈在線粒體中的核糖體,比一般核糖體小,約為55S(35S和25S大、小亞基),稱為胞器或線粒體核體。凡是幼稚的、未分化的細胞、胚胎細胞、培養細胞、腫瘤細胞,它們生長迅速,在胞質中一般具有大量遊離核糖體。真核細胞含有較多的核糖體,每個細胞平均有106~107個,而原核細胞中核糖體較少每個細胞平均只有15×102~18×103個。真核細胞核糖體的沉降係數為80S,大亞基為60S,小亞基為40S。在大亞基中,有大約49種蛋白質,另外有三種rRNA∶28SrRNA、5SrRNA和5.8SrRNA。小亞基含有大約33種蛋白質,一種18S的rRNA。
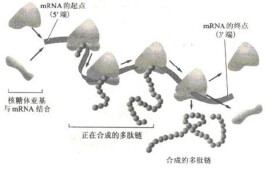
無論哪種核糖體,在執行功能時,即進行蛋白質合成時,常3-5個或幾十個甚至更多聚集並與mRNA結合在一起,由mRNA分子與小亞基凹溝處結合,再與大亞基結合,形成一串,稱為多聚核糖體(遊離多聚核糖體及固著多聚核糖體),Polyribosome或Polysome。mRNA的長短,決定多聚核糖體的多少,可排列成螺紋狀,念珠狀等,多聚核糖體是合成蛋白質的功能團。此時,每一核糖體上均在以mRNA的密碼為模板,翻譯成蛋白質的氨基酸順序。在活細胞中,核糖體的大小亞基,單核糖體和多聚核糖體是處於一種不斷解聚與聚合的動態平衡中,隨功能而變化,執行功能量為多聚核糖體、功能完成後解聚為大、小亞基。
非膜相結構,大小15-20nm,可單個或成群分佈於細胞質中,也可附著在核外膜,內質網上,或存在於線粒體,葉綠體中,用負染色高分辨電鏡觀察,核糖體不是圓形顆粒,而是由大、小二個亞基組成的不規則顆粒。
大亞基側面觀是低面向上的倒圓錐形,底面不是平的,邊緣有三個突起,中央為一凹陷,似沙發的靠背和扶手。小亞基是略帶弧形的長條,一面稍凹陷,一面稍外突,約1/3處有一細縊痕,將其分成大小兩個不等部份。小亞基趴在大亞基上,似沙發上趴了一隻小猴。大小亞基凹陷部位彼此對應相結合,就形成了一個內部空間。此部位可容納mRNA、tRNA及進行氨基酸結合等反應。
此外,在大亞基內有一垂直的通道為中央管,所合成的多肽鏈由此排放,以免受蛋白酶的分解。一般真核細胞中,106-107個/細胞,原核細胞中15-18×103個/細胞,蛋白質合成旺盛的細胞可達1×1012個/細胞。
核糖體的主要成份為蛋白質和rRNA,二者比例在原核細胞中為1.5:1,在真核細胞中為1:1,每個亞基中,以一條或二條高度摺疊的rRNA為骨架,將幾十種蛋白質組織起來,緊密結合,使rRNA大部份圍在內部,小部份露在表面。由於RNA的磷酸基帶負電荷超過了蛋白質帶的正電荷,因而核糖體顯強的負電性,易與陽離子和鹼性染料結合。
單個核糖體上存在四個活性部位,在蛋白質合成中各有專一的識別作用。
1.A部位:氨基酸部位或受位:主要在大亞基上,是接受氨醯基-tRNA的部位。
2.P部位:肽基部位或供位:主要在小亞基上,是釋放tRNA的部位。
4.GTP酶部位:即轉位酶,簡稱g因子,對GTP具有活性,催化肽鍵從供體部位→受體部位。
另外,核糖體上還有許多與起始因子、延長因子、釋放因子以及各種酶相結合的位點。核糖體的大小是以沉降係數S來表示,S數值越大、顆粒越大、分子量越大。原核細胞與真核細胞核糖體的大小亞基是不同的。
50S(大亞基)23S,5SRNAS+原核(70S)34種蛋白質55種蛋白質30S(小亞基)21種蛋白質+16SRNA
真核(80S)60S(大亞基)28S5.8S5SRNA+45種蛋白質78種蛋白質40S(小亞基)33種蛋白質,+18SRNA
抗體是由核糖體合成
(一)蛋白質合成的細胞內定位
核糖體的功能就是將mRNA上的遺傳密碼(核苷酸順序)翻譯成多肽鏈上的氨基酸順序。因此,它是肽鏈的裝配機,即細胞內蛋白質合成的場所,細胞合成的蛋白質可分為兩類:外輸性蛋白和內源性蛋白。
1.外輸性蛋白:主要在固著核糖體上合成,分泌到細胞外發揮作用,如抗體蛋白、蛋白類激素、酶原、唾液等,也能合成部份自身結構蛋白,如膜嵌入蛋白、溶酶體蛋白。
2.內源性蛋白:又稱結構蛋白,是指用於細胞本身或組成自身結構的蛋白質,主要是在遊離核糖體上合成,如紅細胞中的血紅蛋白,肌細胞中的肌纖維蛋白。
(二)蛋白質生物合成的簡要過程
蛋白質生物合成是一個複雜而重要的生命活動,它在細胞中有粗細的結構基礎,進行得十分迅速有效,是依靠分子水平上的嚴密組織和準確控制進行的。
蛋白質合成不僅要有合成的場所,而且還必須有mRNA、tRNA、20種氨基酸原料和一些蛋白質因子及酶。Mg、K+離子等參與,並由ATP、GTP提供能量,合成中mRNA是編碼2合成蛋白質的模板,tRNA是識別密碼子,轉運相應氨基酸的工具。核糖體則是蛋白質的裝配機,它不僅組織了mRNA和rRNA的相互識別,將遺傳密碼翻譯成蛋白質的氨基酸順序,並且控制了多肽鏈的形成,下面看看真核細胞中蛋白質合成的主要步驟,是怎樣在細胞內超微結構水平上進行的。
1.氨基酸的激素和轉運
階段在胞質中進行,氨基酸本身不認識密碼,自己也不會到Ribosome上,須靠tRNA。
氨基酸+tRNA→→氨基醯tRNA複合物
每一種氨基酸均有專一的氨基醯-tRNA合成酶催化,此酶首先激活氨基酸的羥基,使它與特定的tRNA結合,形成氨基醯tRNA複合物。所以,此酶是高度專一的,能識別並反應對應的氨基酸與其tRNA,而tRNA能以反密碼子識別密碼子,將相應的氨基酸轉運到核糖體上合成肽鏈。
2.在多聚核糖體上的mRNA分子上形成多肽鏈
氨基酸在核糖體上的聚合作用,是合成的主要內容,可分為三個步驟:
(1)多肽鏈的起始:mRNA從核到胞質,在起始因子和Mg的作用下,小亞基與mRNA的起始部位結合,甲硫氨醯(蛋氨酸)—tRNA的反密碼子,識別mRNA上的起始密碼AuG(mRNA)互補結合,接著大亞基也結合上去,核糖體上一次可容納二個密碼子。
(2)多肽鏈的延長:第二個密碼對應的氨醯基—tRNA進入核糖體的A位,也稱受位,密碼與反密碼的氫鍵,互補結合。在大亞基上的多肽鏈轉移酶(轉肽酶)作用下,供位(P位)的tRNA攜帶的氨基酸轉移到A位的氨基酸后並與之形成肽鍵(—CO-NH—),tRNA脫離P位並離開P位,重新進入胞質,同時,核糖體沿mRNA往前移動,新的密碼又處於核糖體的A位,與之對應的新氨基醯-tRNA又入A位,轉肽鍵把二肽掛於此氨基酸后形成三肽,ribosome又往前移動,由此漸進漸進,如此反覆循環,就使mRNA上的核苷酸順序轉變為氨基酸的排列順序。
注意:P位(供位):供tRNA;供肽鏈
A位(受位):受氨基酸-tRNA;受肽鏈核苷酸與氨基酸相連繫的橋樑是tRNA。
(3)多肽鏈的終止與釋放:肽鏈的延長不是無限止的,當mRNA上出現終止密碼時(UGA,U氨基酸和UGA),就無對應的氨基酸運入核糖體,肽鏈的合成停止,而被終止因子識別,進入A位,抑制轉肽酶作用,使多肽鏈與tRNA之間水解脫下,順著大亞基中央管全部釋放出,離開核糖體,同時大小亞基與mRNA分離,可再與mRNA起始密碼處結合,也可遊離於胞質中或被降解,mRNA也可被降解。
這是在一個核糖體上氨基酸聚合成肽鏈,每一個核糖體一秒鐘可翻譯40個密碼子形成40個氨基酸肽鍵,其合成肽鏈效率極高。可見,核糖體是肽鏈的裝配機。
合成的若是結構蛋白,則這些多肽便經過某些修飾、剪接后形成四級結構,投入使用,若是輸出蛋白呢?
我們知道分泌蛋白質是先存在於內質網腔中,后經高爾基體排出,胞吐外排,那麼,合成的輸出蛋白是怎樣進入內質網腔的呢?
3.信號學說:Signalhypothesis
與膜結合的核糖體和遊離核糖體在性質上是一樣的,那這種核糖體為什麼會結合到粗面內質網膜上呢?新肽鏈又是怎樣進入RER囊腔的呢?信號學說闡明了固著核糖體上合成蛋白質的特殊性,該學說的基本要點。
(1)分泌蛋白質多肽的合成一開始也在遊離多聚核糖體上,但其mRNA在AUG之後有一段45-90bp的信號順序(密碼),由此能翻譯出15-30個氨基酸的多肽(信號肽)SignalPeptide。這種能合成信號肽的核糖體將成為附著核糖體與內質網結合,不能合成信號肽的為遊離核糖體,仍散佈於胞質中。
(2)近幾年的研究發現,胞質中存在著信號識別顆粒(SignalRecoynitionParticle,SRP),它既能識別露出核糖體之外的信號肽,又能識別RER膜上的SRP受體,只有當核糖體出現信號肽時,SRP才與核糖體的親和力增高。
(3)SRP與核糖體一結合,便以tRNA的構型佔據了核糖體的“A”位,使核糖體的蛋白質合成暫時停止。
(4)SRP-核糖體複合體與RER上的SRP受體結合核糖體則以大亞基結合於RER上的嵌入蛋白(核糖體結合蛋白Ⅰ和Ⅱ),所以SRP受體又稱停泊蛋白(docking蛋白質),SRP與SRP受體結合是暫時的,當核糖體附著於內質網膜后,SRP便離去,核糖體結合蛋白只存在於RER上。
(5)信號肽由疏水性氨基酸構成,當能合成信號肽的核糖體與內質網膜結合后,信號肽便經由內質網膜插入膜腔內,(內質網膜中2-多個識別信號肽的受體蛋白側向移動,集中在一起形成臨時性管道與中央管相連接),而先前處於暫停狀態的蛋白質合成活動又重新開始。進入內質網腔的信號肽將與之相連的新生肽鏈引入內質網腔。信號肽便被位於內質網內表面的信號肽酶切掉,核糖體繼續合成肽鏈,肽鏈不斷延長,並在內質網腔中保護不被破壞並在網腔中形成具有一定空間構型的蛋白質,當合成終止,受體蛋白重新分散,肽鏈從核糖體脫下,核糖體大小亞基離開,所以,固著核糖體與RER的結合不是結構性的,而是特異性、暫時性、功能性的。
所以,如信號順序發生改變,所合成的信號肽不能被受體識別,核糖體就結合不到膜上。
電鏡下,多聚核糖體的解聚和粗面內質網的脫粒都可看作是蛋白質合成降低或停止的一個形態指標。
多聚核糖體的解聚:是指多聚核糖體分散為單體,失去正常有規律排列,孤立地分散在胞質中或附在粗面內質網膜上。一般認為,遊離多聚核糖體的解聚將伴隨著內源性蛋白質生成的減少。脫粒是指粗面內質網上的核糖體脫落下來,分佈稀疏,散在胞質中,RER上解聚和脫離將伴隨外輸入蛋白合成。
正常情況下,蛋白質合成旺盛時,細胞質中充滿多聚核糖體,RER上附有許多念珠線狀和螺旋狀的多原核糖體,當細胞處於有絲分裂階段時,蛋白質合成明顯下降,多聚核糖體也出現解聚原C,逐漸為分散孤立的單體所代替。
另外,一些藥物,致癌物可直接抑制蛋白質合成的不同階段,有些抗苔素,如鏈黴素、氯黴素、紅黴素等對原核與真核生物的敏感性不同,能直接抑制細菌核糖體上蛋白質的合成作用。有的抑制在起始階段,有的抑制肽鏈延長和終止階段,有的阻止小亞基與mRNA的起始結合,四環素抑制氨基醯-tRNA的結合和終止因子,氯黴素抑制轉肽酶,阻止肽鏈形成,紅黴素抑制轉位酶,不能相應移位進入新密碼。所以,抗苔素的抗苔作用就是干擾了細苔蛋白合成而抑制細苔生長來起作用的。
1、核糖體的結構特點——暫時的、功能的動態結構。
2、核糖體種類與合成蛋白質種類的關係。
3、留下思考題:核糖體合成的蛋白質之去向(與內膜系統聯繫)。
核糖體不是由生物膜構成的,它是由蛋白質和RNA構成的複合體。由大小兩個亞基組成。核糖體是蛋白質合成的場所。附著在內質網上的核糖體合成的蛋白質主要有兩類:一類是分泌蛋白,通過內質網運輸到高爾基體,經加工包裝后被分泌到細胞外;另一類是排列到質膜內的蛋白質。遊離的核糖體合成的蛋白質一般是分佈到細胞質基質中的蛋白質,如分佈於細胞質基質中的酶等
基本信息
- 中文名
- 聚核糖體
- 外文名
- ribosome
- 主要構成
- rRNA和蛋白質
- 別名
- 核蛋白體