細胞質遺傳
細胞質遺傳
細胞質遺傳(cytoplasmic inheritance)是指子代的性狀由細胞質內的基因所控制的,是細胞質內的基因所控制的遺傳現象和遺傳規律。
細胞質基因:線粒體、葉綠體中的DNA上和細胞質粒上的基因。
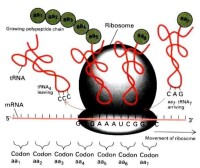
細胞質遺傳現象表明,細胞質內具有控制某些性狀的遺傳物質——細胞質基因(簡稱質基因)。但是,科學家用電子顯微鏡觀察,在細胞質內並沒有找到像染色體一樣的結構。1962年,科學家裡斯(Ris)和普蘭特(Plant)等用電子顯微鏡觀察衣藻、玉米等植物的超薄切片,發現在葉綠體的基質中有長度為20.5nm左右的細纖維存在。用DNA酶處理,這種細纖維就消失。由此證明,這種細纖維結構就是葉綠體DNA。
細胞質基因:線粒體、葉綠體中的DNA上和細胞質粒上的基因。
細胞質遺傳現象表明,細胞質內具有控制某些性狀的遺傳物質——細胞質基因(簡稱質基因)。但是,科學家用電子顯微鏡觀察,在細胞質內並沒有找到像染色體一樣的結構。1962年,科學家裡斯(Ris)和普蘭特(Plant)等用電子顯微鏡觀察衣藻、玉米等植物的超薄切片,發現在葉綠體的基質中有長度為20.5nm左右的細纖維存在。用DNA酶處理,這種細纖維就消失。由此證明,這種細纖維結構就是葉綠體DNA。
細胞質遺傳
細胞質遺傳
②雜交後代不出現一定的分離比。
(2)原因
①受精卵中的細胞質幾乎全部來自卵細胞;
②減數分裂時,細胞質中的遺傳物質隨機不均等分配。
質粒:原核、細菌、小環DNA。鬆弛型和嚴緊型2類。
線粒體基因:mtDNA,線狀、環狀,能單獨複製,同時受核基因控制。哺乳動物:無內含子,有重疊基因突變率高。
葉綠體基因:ctDNA,環狀,可自主複製,也受核基因控制。
卡巴粒:草履蟲體內細胞質顆粒
細胞質遺傳:子代的性狀是由細胞質內的基因所控制的遺傳現象。也叫母系遺傳、核外遺傳、細胞質遺傳、母體遺傳、非孟德爾式遺傳。
(一)高等植物葉綠體的遺傳
有幾種高等植物有綠白斑植株,如紫茉莉、藏報春、加荊介等。1901年柯倫斯在紫茉莉中發現有一種花斑植株,著生綠色,白色和花斑三種枝條。在顯微鏡下觀察,綠葉和花斑葉的綠色部分其細胞中均含正常的葉綠體,而白色或花斑葉的白色部分,細胞中缺乏正常的葉綠體,是一些敗育的無色顆粒。
他分別以這三種枝條上的花作母本,用三種枝條上的花粉分別授給上述每個作為母本的花上,雜交後代的表現:完全取決於母本,而與花粉來自哪一種枝條無關。
接受花粉的枝條 提供花粉的枝條 雜種表現
細胞質遺傳
花 斑
白 色
綠 色 綠 色 綠 色
花 斑
白 色
花 斑 綠 色 白、綠、花斑
花 斑
上述雜交結果說明:決定性狀表現的遺傳基礎就在細胞質中,通過許多科學家的不同實驗,都證明了細胞質的葉綠體含DNA,是雙螺旋線性或環狀裸露DNA分子,能自我複製,穩定遺傳。葉綠體的遺傳不僅受細胞質基因的控制,還於核基因有關。
(二)真菌類的線粒體遺傳
1、酵母菌小菌落的遺傳:啤酒酵母屬於子囊菌,它在有性生殖時,不同交配型相結合形成的二倍體合子。酵母有一種“小菌落”個體。這種類型經培養后只能產生小菌落。
如果把小菌落酵母同正常個體交配,則產生正常的二倍體合子。經減數分裂產生單倍體後代也表現正常,不再分離小菌落。這表明小菌落性狀的遺傳與細胞質有關,而且這種交後代,4個子囊孢子有2個是a+,另兩個是a-,交配型基因a+和a-仍然按預期的孟德爾比例時行分離,而小菌落性狀沒有象核基因那樣發生重組和分離,說明這個性狀與核基因無關。
進一步研究發現,小菌落酶母細胞內,缺少細胞色素a和b,還缺少細胞色素氧化酶,這些酶類,存在於細粒體中,表明這種小菌落的變異與線粒體的基因組變異有關。
2、鏈孢霉緩慢生長型的遺傳:鏈孢霉有一種緩慢生長突變型,呼吸弱,生長慢,這是由於線粒體結構和功能不正常,細胞色素氧化酶缺少,氧化作用降低,生長緩慢。
進行有性生殖時,正反交結果不同:雜交後代不出現一定的分離,表現非孟德爾式遺傳。如果突變型的單倍體核(♂)和野生型的受精絲中的單倍體核(♀)結合(雜交),得到的後代全是野生型,反之全是突變型。
核質互作控制性狀的實例:
(一)細胞質基因受核基因的控制
1、玉米埃型條斑的遺傳:玉米第7染色體上有一個條紋基因ij,當起處於隱性純合時(ijij),能引起質體突變率增加,使正常的質體突變為敗育的質體,不能全部形成葉綠素,表現出白色和綠色相間的條斑性狀的植株或是白化苗不能成活。
當條斑為母本與正常株IjIj為父本雜交時,其F1(Ijij)表型由種:綠色苗、條斑苗和白化苗。表明受這對基因控制的突變體是通過母本細胞質遺傳的。如果反交,F1基因型與正交F1相同,但因母本細胞核中無ijij純合基因,不能引起變異,因此雜種只表現綠苗的一種類型,說明質體變異是受核基因控制的。
2、草履蟲放毒型的遺傳:草履蟲是單細胞原生動物,已知有兩個品系,一個叫做放毒型(能產生草履蟲素,殺死其他品系的草履蟲而對自身無害),另一個叫敏感型(不能產生毒素,而且易被草履蟲素所殺)。
放毒型草履蟲為什麼會產生草履蟲素呢?是由於兩個因素的相互作用而共同決定的:其一是在它的細胞質中,大約含有幾百個卡巴粒(推測是一種含有噬菌體的溶源性細菌),這種卡巴粒含有DNA,並能自我複製和發生變異,表明在卡巴粒中含有質基因。其二是它的細胞核中含有一個顯性基因K,只有當卡巴粒和顯性基因K同時存在於一個草履蟲內時,它才產生毒素,並是穩定放毒型。
草履蟲有一個大核,主要負責營養,多倍性的,還有兩個小核,是二倍性的,主要與遺傳有關。草履蟲的生殖方式有2種:無性生殖和有性生殖。
無性生殖:由一個個體通過細胞分裂,成為兩個個體,基因型仍跟原來的個體一樣。
有性生殖也稱接合生殖,兩個體接合,相互交換小核,此過程中大核消失,小核經減數分裂后相互交換。另外草履蟲也可以自體受精。
當放毒型品系與敏感型品系雜交,如果接合時間長,相互交換了小核,同時也交換了細胞質,則都變成了放毒型,如果接合時間短,只交換了小核,沒有交換細胞質,則交換后,放毒型仍為放毒型,敏感型也為敏感型。兩種接合生殖情況,結果不同。可見,放毒性狀的表現必須是:
KK+卡巴粒→放毒型
kk+卡巴粒
kk無卡巴粒 敏感型
KK卡巴粒
(二)細胞質對核基因作用的調節
1、細胞質對基因載體—染色體的調節
植物雄性不育的遺傳
2、細胞質對X染色體上基因的調節作用
哺乳動物性染色體♀XX,♂XY。X染色體上含有很多與性別無關的伴性基因。按理說,這樣的基因♀性有兩套,♂性只有1套,♀性基因產物也應是♂性基因產物地倍,可事實上並不是這樣,二者產物基本相等。
因為♀性的兩個X染色體,在間期核中表現“異固縮現象”,即屬於異染色質,染色深,處於失活狀態。至於哪個細胞中哪條染色體失活是隨機的。
婦產科可以根據胎兒羊水中的胎兒脫落細胞染色,判斷是男孩還是女孩。
(三)個體發育中質基因與核基因的關係
1、細胞質基因在個體發育中的作用是必須的,不可缺少的。
2、細胞核基因在個體發育中的起主導作用。
3、核質互作辨證統一的關係
生物遺傳性狀中,有兩個主要的遺傳系統—核遺傳和質遺傳,它們除了有相對的獨立性,核基因起主導作用外,但又密切的聯繫,互相協調,在遺傳上綜合地發揮作用。
四、持續飾變
環境引起的表現型改變通過母親細胞質而連續傳遞幾代,變異性逐漸減少,最終消失的遺傳現象。
介於母性影響和細胞質遺傳之間。不能隔代遺傳,無論在後代中怎樣選擇,最終性狀消失。
例如,用水化氯醛處理四季豆,葉子產生畸形。F1代73%,F2代67%,F3代47%,F4代52%,F5代8%,F6代4%,F7代0%。
細胞質遺傳
指母親的基因型僅影響子代個體的幼齡期。
例如麥粉蛾(麵粉蛾)的色素遺傳中的母性影響。
野生型:幼蟲皮膚有色,成蟲複眼深褐色。
突變型:幼蟲皮膚無色,成蟲複眼紅色(缺乏犬尿素)。
P 有色AA×aa無色→F1 Aa 有色
↙ ↘
♀aa ×Aa♂ ♀Aa×aa♂
↓ ↓
1/2Aa 1/2aa 1/2Aa 1/2aa
幼蟲:有色 無色 有色 有色
成蟲複眼:褐色 紅色 褐色 紅色
核基因AA→犬尿素(細胞質)→色素
指母親的基因型對子代個體的影響是持久的,影響子代個體的終生。
例如:錐實螺的外殼旋轉方向的遺傳就是母性影響一個比較典型的例證。錐實螺♀♂同體,繁殖時異體受精,但是如果是單個飼養,就自體受精。
錐實螺的外殼旋轉方向可以是右旋的,也可以是左旋的。判斷的方法:使螺殼的開口朝向自己,如果開口在右邊即右旋,如果開口在左邊即左旋。
左旋、右旋由一對基因控制,右旋D對左旋d是顯性。右旋與左旋交配,依雌性的旋向不同,結果也不同。 P ♀DD×♂dd P ♀dd×♂ DD
右↓左 左↓右
F1 Dd右 F1 Dd左
↓自交 ↓自交
F2 1DD 2Dd 1dd全右 F2 1DD 2Dd 1dd全右
↓自交 ↓自交
F3 右旋:左旋=3:1 F3 右旋:左旋=3:1
上述這種現象如何解釋呢?研究表明,螺類的受精卵是通過螺旋式的卵裂開始幼體發育的,未來的螺殼旋向決定於最初兩次卵裂中紡錘體的方向。紡錘體的方向又取決與卵細胞質的特性,而卵細胞質的特性歸根到底又取決於母本的基因型。這樣就形成了母本的基因型控制子代的表現型的現象(只控制子代)。雖然母本細胞質特徵決定了子代的表型,但實質是核基因的遺傳仍符合孟德爾式的遺傳方式。
植物雄性不育的遺傳
定義
不育:一個個體不能產生有功能的配子或不能產生在一定條件下能夠存活的合子的現象。
雄性不育性:植株不能產生正常的花藥、花粉或雄配子,但它的雌蕊正常,能接受正常花粉而受精結實。
類型
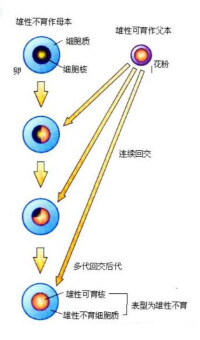
1、細胞質雄性不育(質不育型):由細胞質基因控制,一般不受父本基因型影響。應用價值不大。
如:質不育型♀×♂正常品系
↓
F1 全部雄性不育♀×♂正常品系
↓多代
全部雄性不育
(1)不受光溫影響的核雄性不育:與光溫影響無關。
(2)光溫敏核雄性不育:受光和溫度影響。高溫或長日——不育;適溫短日——可育。
3、核質互作雄性不育型:也叫質核互作型,核基因和細胞質基因共同作用控制的雄性不育類型。即細胞質雄性不育基因S,核內又具有純合的核不育基因rfrf,才能表現雄性不育,二者缺一不可。細胞質可育基因(N),核可育基因RfRf。
機制
還不是很清楚,只是有一些關於核質互作不育的機理假說。
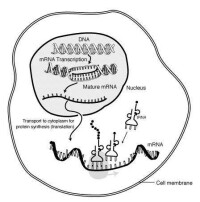
1、質核互補控制假說:認為細胞質不育基因位於線粒體體內。在胞質正常的情況下(N),線粒體DNA可攜帶可育的遺傳信息,經轉錄合成正常的mRNA,繼而在線粒體核糖體上合成各種蛋白質,從而保證雄蕊發育過程中,全部代謝活動正常進行,最終導致形成結構功能正常的花粉。
當線粒體DNA發生變異后,只要核基因中有Rf基因,就可能轉錄出mRNA,合成花粉發育用的蛋白質等,如果核基因rf,因無補償作用而表現為花粉不育。如果N與Rf同時存在,N就起調節基因作用。
2、病毒危害假說:認為造成不育的原因,可能是由於病毒侵害造成的,這種病毒侵染敏感菌株(rfrf)后,不危害2倍體宿主細胞,但卻能殺死單倍體的花粉,而Rf能給予花粉以一種抗性,來抵抗病毒的有害作用。
應用
主要在雜種優勢上的應用,雜交母本獲得雄性不育后,就可以節省大面積制種時的去雄勞動量。並保證雜交種子的純度。但在應用時,必須三系配套,具備不育系、保持系和恢復系。
(一)三系 的培育(玉米為例)
1、不育系和保持系的培育:
雄性不育系基因型:(S)rfrf

植物雄性不育的遺傳
保持系:能夠保持雄性不育系特性的品系或品種。保持系本身是可育的,當它與不育系雜交,雜交後代仍能保持不育系的不育性。只作雜交父本用。簡稱B系。
(S)rfrf×(N)rfrf→(S)rfrf
↓自交
(N)rfrf
2、恢復系的培育
恢復系(N)RfRf:能使雄性不育系的後代恢復其育性的品種或品系。恢復系與雄性不育系雜交,就能獲得可育的雜交種,以供大田生產之用。作雜交父本,簡稱C系。
恢復系Ä→恢復系,進行繁殖。
利用方法
※二區三系制種法
1、第一區:是不育系和保持系繁殖區(隔離區),在此區交替種植不育系和保持系,二者在開花時,保持系給不育系提供花粉雜交,同時也自交,在保持系植株上收穫保持系。
2、第二區:制種區,(雜種制種隔離區)交替種植不育系和恢復系。恢復系給不育系提供花粉,生產雜交種,供大田使用。而恢復系植株自花授粉繁殖恢復系的種子。在實際生產中,由於作物種類不同,方法也大同小異,但必須與明顯的雜種優勢,才能值得配製三系。
※二系法
1973年,石明松在晚粳農墾58中發現“湖北光敏核不育水稻—農墾58S”。在長日照下不育,短日照下可育→可將不育系和保持系合二為一→提出生產雜交種子”二系法”。
光敏核不育系
細胞質遺傳
↓ 自交 ↓ ↓自交
光敏核不育系 F1雜交種 恢復系
系統影響
1、不育系的不育度要高,應接近100%,
2、恢復系的花粉量要大,恢復力要強,至少要達到85%的恢復度,
3、不育系與恢復系間的雜種優勢要強。
