染色體外遺傳
染色體外遺傳
染色體外遺傳
染色體外遺傳
染色體外遺傳
1924 年德國學者 R.von 韋特施泰因報道了葫蘆蘚(Furnaria)中的母體遺傳現象,並提出細胞質基因組這一名詞,用來概括細胞質中的全部遺傳物質。
P.米凱利斯從 20年代開始對柳葉菜屬(Epilobium)植物連續進行了二、三十年的研究,分析了若干種性狀的母體遺傳現象。在這一段時期中還出現了一些有關細胞質遺傳的名詞。例如1937年日本學者今井提出質體基因,用來指葉綠體中的遺傳因子;1939年英國細胞學家C.D.達林頓提出細胞質基因,用來泛指細胞質中的獨立自主的遺傳因子。1937年法國學者P.萊里蒂埃報道了果蠅對於二氧化碳敏感性狀的非孟德爾式遺傳現象。隨後證實這種現象是果蠅被σ病毒感染所致。
1943年美國學者M.M.羅茲在玉米中報道一種白色條紋葉──埃型條紋性狀的非孟德爾式遺傳現象。後來知道埃型條紋性狀來源於核基因突變,這是核基因和細胞質基因相互作用的第一個例子。
40年代中微生物遺傳學研究廣泛開展以後又陸續在酵母菌、脈孢菌等微生物中發現染色體外遺傳現象。1943年美國學者T.M.索恩本在雙小核草履蟲中發現放毒性的遺傳屬於非染色體遺傳,而且證明這是由一種和草履蟲共生的微生物卡巴帶來的性狀。1949年法國學者B.埃弗呂西等在啤酒酵母中發現了小菌落突變型 (petite,p-,它的線粒體中缺少某些細胞色素成分。小菌落突變型有染色體外遺傳和染色體遺傳兩類,前者稱為營養型小菌落,後者稱為分離型小菌落。這兩類突變型的存在說明線粒體的形成同時受核基因和線粒體本身的控制。1952年美國學者J.萊德伯格繼 50年代初大腸桿菌中的F因子(見細菌接合)和λ噬菌體中的溶源性(見轉導)的發現之後,提出了質粒這一名詞,用來指染色體外的全部獨立自主的遺傳因子,包括σ因子等共生生物、線粒體和葉綠體等細胞器以及F因子這一類單純的DNA分子。不過現在質粒這一名詞已用來專指最後一類遺傳因子了。1963~1964年分別確證了線粒體和葉綠體中存在著DNA,從此以後染色體外遺傳的研究已逐漸成為分子遺傳學中的重要領域。
染色體外遺傳
多數微生物是單倍體(見染色體倍性),染色體基因在雜交子代中按 1:1分離。例如啤酒酵母的分離性小菌落和野生型的雜交子代中雙親的類型按 1:1分離,這說明分離性小菌落的遺傳屬於染色體遺傳範疇。可是表型同屬於呼吸缺陷的營養性小菌落和野生型的雜交子代全部都屬於野生型,說明營養性小菌落的遺傳屬於染色體外遺傳。脈孢菌的po突變型除了表現正反交結果不相同現象以外,實際上也表現不分離現象。
性狀隨著染色體以外的細胞質成分的轉移而改變,按照生物的特性,可以通過不同的途徑把一個生物的細胞質轉移到另一生物中,如果后一生物因此而獲得前一生物的某些性狀,說明這一性狀的遺傳方式屬染色體外遺傳。細胞質可以通過多種方式轉移。
在某些種類的果蠅中有一種稱為性比的因子。它使子代中只出現雌性個體。把具有這一性狀的果蠅的體液注射到正常的雌蠅體內,可以使後者變為只產雌蠅。黑腹果蠅(Drosophila melanogaster)中的另一性狀──對二氧化碳敏感也屬於同一類型。關於粗糙鏈孢黴菌的po性狀,也曾用注射去核的細胞質的方法證明它的遺傳方式屬染色體外遺傳。
大腸桿菌(Escherichia coli)可以通過細菌接合在不發生染色體轉移的情況下轉移它的 F因子,而且使接受F因子的細菌的後代都具有F因子,說明F因子的遺傳屬染色體外遺傳。同樣證實屬染色體外遺傳的還有草履蟲的放毒性、大腸桿菌以及其他一些細菌的抗藥性等。
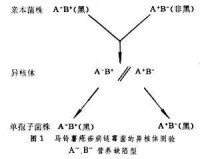
放線菌的菌絲細胞很容易聯接在一起。如兩個核基因有區別的菌株的菌絲相互聯接,從聯接處長出的菌絲中便有兩種核,這樣的菌絲體稱為異核體。如果這是一種產生單核分生孢子的生物,那麼在形成分生孢子時異核體的兩種細胞核分別出現在不同的分生孢子中,但是已經混合的細胞質則不再分開。按照核基因標記分離兩種單孢子菌株,如發現有性狀轉移,就說明這一性狀的轉移和細胞質的混合有關,因而屬於染色體外遺傳。馬鈴薯瘡痂病鏈黴菌 (Streptomycesscabies)黑色素的遺傳可以說明這一原理(圖1)。染色體外遺傳
通過多次和雄性親本回交,最後可以得到細胞質來自一個品系而染色體則幾乎全部來自另一品系的雜交子代。如果雌性親本的某些性狀並不由於通過多次回交后染色體被取代而改變,便可以認為這些性狀的遺傳屬於染色體外遺傳。在柳葉菜和脈孢菌中,都曾用此法證明了一些性狀的遺傳屬於染色體外遺傳(見雜交)。
染色體外遺傳
某些性狀可以由於一些環境因素的處理而被消除,這也是判斷染色體外遺傳的一種依據。例如吖啶類染料可以使一些抗藥性細菌變為敏感的細菌;加速草履蟲放毒品系的分裂速度可以使它變為非放毒品系等。染色體基因突變型可以通過回復突變而成為原來的野生型,可是被消除的性狀不再重新出現,說明這些性狀的遺傳依賴於染色體外的遺傳因子。
染色體外遺傳是染色體外的遺傳因子(細胞器、內共生生物和質粒)傳遞的結果。
共生生物在真核生物和原核生物中都有發現。質粒雖然最初是在原核生物的細菌中發現的,而且大量的質粒都是在細菌中發現,但在低等的真核生物如酵母菌中也有發現。內共生體和質粒雖然也編碼某些蛋白質,並且控制宿主細胞的某些遺傳性狀,但都不是生命活動所必需的。
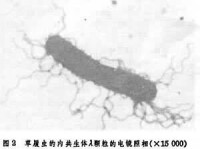
草履蟲的放毒性狀是由於放毒品系細胞中有一種卡巴(k)顆粒。卡巴粒長約1~5微米,外面有雙層膜,革蘭氏染色陰性,富爾根染色陽性,含有DNA、RNA以及一些酶系和細胞色素。每一細胞中的卡巴粒數目可以少到幾個,多到幾百個。卡巴粒還可以發生突變。雖然卡巴粒的離體培養還沒有成功,可是一些和它相似的顆粒如萊姆達(λ)和繆(μ)都可以離體培養。已經公認它們是一類和細菌相類似的共生生物(圖2)。
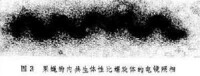
同樣地已經證明果蠅的性比因子是一種螺旋體(圖3)。果蠅的二氧化碳敏感因子是一種病毒,呈炮彈形,和水泡性口膜炎病毒很相似。
見質粒。
酵母菌的小菌落突變型就是線粒體發生缺陷的結果。
酵母菌中呈現染色體外遺傳的性狀除了小菌落突變型以外,還有氯黴素抗性(chloramphenicol resistance,cap)、紅黴素抗性(erythromycin resistance,ery)、寡黴素抗性(oligomycin resistance,oli)、巴龍黴素抗性(paromomycinresistance,par)等。線粒體遺傳學研究主要通過三個方面進行:
分析方法在原則上和測定減數分裂中基因重組頻率的基因定位方法相同。
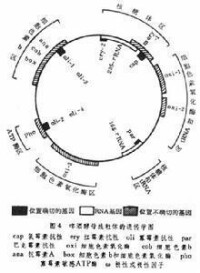
小菌落突變型(p-)可以通過溴化乙錠(EB)等誘發,這樣產生的小菌落突變型常常是線粒體DNA缺失的結果。鄰接的基因常常同時缺失,所以通過許多p-菌株的抗性基因或其他基因的共缺失的分析,可以測定各個基因的相對位置。分子雜交和限制性核酸內切酶物理圖譜分析 結合基因定位和通過分子雜交和限制圖譜等研究,可以初步畫出啤酒酵母的線粒體的遺傳學圖(圖4)。
啤酒酵母的線粒體遺傳學研究中還發現類似於大腸桿菌的性別(見細菌接合)的所謂極性現象(不同的極性或極性因子用ω+和ω-表示)。此外,酵母菌的線粒體遺傳學研究中,還有兩項具有更廣泛的生物學意義的課題。一是斷裂基因如細胞色素b氧化酶基因(cytochromeb oxi-dase,box)的研究,發現這一基因的內含子編碼的所謂成熟酶催化切除mRNA中的內含子成分的反應。另一是關於線粒體的遺傳密碼的研究,這一研究提供了關於密碼進化的有力證據(見分子進化)。
最早發現的染色體外遺傳是高等植物的葉綠體的遺傳。目前葉綠體的遺傳學分析仍以萊因哈德衣藻(Chlamydomonas reinhardi)最為詳盡,它的研究從1954年美國學者R.塞傑取得呈現不分離現象的抗鏈黴素突變型Sr-500開始。葉綠體的遺傳學分析主要通過基因重組分析和純合化分析。純合化分析的原理和利用體細胞重組進行基因定位的原理相同,不過在體細胞重組分析中,基因離著絲粒愈遠則純合化頻率愈高,而在這裡則假定環狀的葉綠體 DNA以某一特定部位附著在葉綠體的膜上,任何基因離這一部位愈遠則純合化頻率愈高或進程愈快。通過這些研究並結合分子雜交等方法的運用,繪出了環狀葉綠體基因的遺傳學圖(圖5)。
關於染色體外遺傳因子和染色體基因的的相互作用關係的研究主要依靠下列幾種方法:①遺傳方式的分析。玉米的白色條紋葉性狀的出現是基因突變的結果,可是它的遺傳卻是非孟德爾式的。酵母菌的小菌落突變型有分離性的(即染色體的)和營養性的(即染色體外的)兩類。草履蟲的卡巴顆粒的遺傳方式屬於染色體外遺傳,可是核基因K的存在對於保持染色體外的卡巴顆粒卻是必要的。這些都說明保持這些遺傳性狀都需要核基因和細胞質基因的相互作用。②分子雜交。一個RNA分子只能和轉錄它的這部分DNA進行分子雜交。因此通過DNA-RNA分子雜交,可以測定編碼某一細胞器成分的基因是在這一細胞器的 DNA分子上還是在染色體上。例如,分子雜交結果說明梨形四膜蟲(Tetrahymenapyriformis)的一部分線粒體 tRNA由染色體基因所編碼。③蛋白質合成分析。例如已經知道啤酒酵母的細胞色素C氧化酶是在線粒體的內膜上,由三個大亞基和四個小亞基構成。離體的酵母菌的線粒體能合成三個大亞基而不能合成四個小亞基,說明前者由線粒體基因編碼而後者並不由它編碼。此外,已經知道抑製劑亞胺環己酮抑制細胞質中的蛋白質合成而紅黴素則抑制線粒體中的蛋白質合成。在紅黴素存在的情況下酵母菌的細胞色素氧化酶三個大亞基的合成被抑制,而亞胺環己酮則不。上述事實都說明同一線粒體的一個部分為線粒體基因編碼,而另一部分則為染色體基因編碼。
已經知道線粒體中由染色體基因和線粒體基因共同參與合成的成分至少有tRNA、腺苷三磷酸酶、細胞色素氧化酶、細胞色素b;葉綠體中染色體基因和葉綠體基因共同參與合成的至少有核糖體蛋白質、tRNA、葉綠體的外膜以及片層結構和光合作用酶系Ⅰ、Ⅱ。對煙草葉綠體中核酮糖磷酸羧化酶合成的控制也曾進行比較深入的研究。
染色體基因和染色體外基因的相互作用還涉及細胞器的裝配問題,在衣藻中發現了不少於20個影響線粒體核糖體裝配的基因,而且其中的 7或 8個是在染色體上。它們中的一部分突變型不能裝配核糖體大亞基,一部分不能裝配小亞基,另外一部分不能裝配大小兩種亞基。葉綠體的形成同樣是一個極為複雜的過程,在大麥中曾發現86個染色體基因和葉綠體的形成有關。
細胞器的形成還受環境的影響,在缺氧條件下生長的酵母菌形成原線粒體。同樣,在無光條件下植物只形成原葉綠體,在有光條件下原葉綠體又轉變成為葉綠體。細胞器的裝配機制和它們的雙重控制機制都有待於深入的研究。
早在1890年德國組織學家R.阿爾特曼便認為線粒體來自共生的細菌,1905~1910年K.C.梅列日科夫斯基同樣認為葉綠體來自共生的生物。在細胞器遺傳的研究取得迅速發展以後,1970年L.馬古利斯提出了更為完整的共生假設,認為在原始的缺氧環境中首先出現了厭氧的原核生物。隨著營光合作用生物的出現,地球上出現了好氧的原核生物和原真核生物,這些原真核生物不具備呼吸酶系。以後好氧原核生物在原真核生物中進行共生,共生體逐漸發展成為好氧的真核生物;營光合作用的原核生物在真核生物中進行共生,共生體逐漸發展成為營光合作用的真核生物。
1972~1975年R.A.拉夫和H.J.馬勒提出非共生進化學說,認為原始的真核生物具備呼吸酶系,而且這些酶系和細菌一樣是在細胞膜上。當細胞變大時,膜向裡面摺疊,並且終於脫離細胞,這些膜把細胞中的帶有某些染色體基因的質粒包裹起來而成為線粒體。另外一些假設並不假定先有共生生物或質粒的存在。例如L.賴恩德斯在1975年提出假說,認為線粒體中的DNA來源於原核細胞本身,被它本身的膜所包被起來,然後通過演變而成為線粒體DNA。
雖然每一種假設的提出都根據了一些事實,但是同樣也都面臨著一些難以解釋的現象。細胞器遺傳本身就是個複雜的過程,它們的演化問題也只能留待今後再作結論。
細胞質遺傳研究的最重要的實踐應用,是雄性不育(見不親和性和雜種優勢)。

基本信息
- 中文名
- 染色體外遺傳
- 外文名
- extrachromosomal inheritance
- 別名
- 核外遺傳
- 類型
- 線粒體遺傳、葉綠體遺傳
- 遺傳特點
- 自主複製