過氧化物酶體
1954年J.Rhodin發現的細胞器
過氧化物酶體由J. Rhodin(1954年)首次在鼠腎小管上皮細胞中發現。是一種具有異質性的細胞器,在不同生物及不同發育階段有所不同。直徑約0.2~1.5um,通常為0.5um,呈圓形,橢圓形或啞呤形不等,由單層膜圍繞而成。共同特點是內含一至多種依賴黃素(flavin)的氧化酶和過氧化氫酶(標誌酶),已發現40多種氧化酶,如L-氨基酸氧化酶,D-氨基酸氧化酶等等,其中尿酸氧化酶(urate oxidase)的含量極高,以至於在有些種類形成酶結晶構成的核心。左圖示:中央具有尿酸氧化酶形成的晶體狀核心
過氧化物酶體(peroxisome)又稱微體(microbody),過氧化物酶體在1954年被發現時, 由於不知道這種顆粒的功能,將它稱為微體(microbody)。過氧化物酶體(peroxisome)是一種細胞器,存在於一切真核細胞內,含有約40餘種氧化酶和觸酶,主要功能是催化脂肪酸的β-氧化,將極長鏈脂肪酸(very long chain fattyacid,VLCFA)分解為短鏈脂肪酸。過氧化物酶體是由一層單位膜包裹的囊泡, 直徑約為0.5~1.0μm, 通常比線粒體小。過氧化物酶體普遍存在於真核生物的各類細胞 中,但在肝細胞和腎細胞中數量特別多。過氧化物酶體含有豐富的酶類,主要是氧化酶,過氧化氫酶和過氧化物酶。氧化酶可作用於不同的底物,其共同特徵是氧化底物的同時,將氧還原成過氧化氫。過氧化物酶體的標誌酶是過氧化氫酶,它的作用主要是將過氧化氫(H2O2, Hydrogen Peroxide)水解。過氧化氫(H2O2)是氧化酶催化的氧化還原反應中產生的細胞毒性物質,氧化酶和過氧化氫酶都存在於過氧化物酶體中,從而對細胞起保護作用。
人們早期認為過氧化酶體的發生與溶酶體類似,但現有證據表明,過氧化酶體的發生與線粒體或葉綠體類似,但在過氧化酶體中不含DNA,組成其的蛋白都在細胞核中編碼,在細胞質基質中產生,再通過信號分選進入過氧化酶體。現在已知的該細胞器的發生有兩種途徑:一是成熟的過氧化酶體經分裂增殖產生子代細胞器;另一種是細胞內的重新發生;這個過程包括三個階段的裝配過程:
Ⅰ過氧化酶體的裝配起始於細胞的內質網,也就是由內質網出芽生成前體膜泡,然後一些過氧化酶體的膜蛋白摻入,形成過氧化酶體雛形(peroxisomal ghost),其中Pex19蛋白作為過氧化酶體膜蛋白靶向序列的胞質受體而發揮作用,簡而言之就是在一定程度上和膜蛋白如PMP70等結合“引導”其到膜上。而Pex3和Pex16輔助過氧化酶體膜蛋白正確插入形成新的前體膜泡。在所有過氧化酶體膜蛋白都插入后,形成過氧化酶體雛形。
Ⅱ具有PTS1和PTS2分選信號的基質蛋白,他們分別以Pex5和Pex7蛋白作為胞質受體,各自與其結合后再與膜受體Pex14結合,在蛋白複合物Pex10、Pex12和Pex2的介導下完成基質蛋白的輸入,形成成熟的過氧化酶體。
Ⅲ成熟的過氧化酶體經分裂產生子代過氧化酶體,分裂過程依賴於Pex11蛋白。
功能:
(1)使毒性物質失活
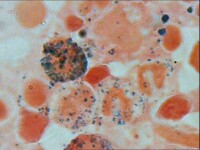
過氧化物酶體
(2)對氧濃度的調節作用
過氧化物酶體與線粒體對氧的敏感性是不一樣的,線粒體氧化所需的最佳氧濃度為2%左右,增加氧濃度,並不提高線粒體的氧化能力。過氧化物酶體的氧化率是隨氧張力增強而成正比地提高。因此,在低濃度氧的條件下,線粒體利用氧的能力比過氧化物酶體強,但在高濃度氧的情況下,過氧化物酶體的氧化反應佔主導地位,這種特性使過氧化物酶體具有使細胞免受高濃度氧的毒性作用。
(3)脂肪酸的氧化
動物組織中大約有25~50%的脂肪酸是在過氧化物酶體中氧化的,其他則是在線粒體中氧化的。另外,由於過氧化物酶體中有與磷脂合成相關的酶,所以過氧化物酶體也參與脂的合成。
(4)含氮物質的代謝
在大多數動物細胞中,尿酸氧化酶(urate oxidase)對於尿酸的氧化是必需的。尿酸是核苷酸和某些蛋白質降解代謝的產物,尿酸氧化酶可將這種代謝廢物進一步氧化去除。另外,過氧化物酶體還參與其他的氮代謝,如轉氨酶(aminotransferase)催化氨基的轉移。
反應:
各類氧化酶的共性是將底物氧化后,生成過氧化氫。
RH2+O2→R+H2O2
過氧化氫酶又可以利用過氧化氫,將其它底物(如醛、醇、酚)氧化。
R′H2+H2O2→R′+2H2O 此外當細胞中的H2O2過剩時,過氧化氫酶亦可催化以下反應:
2H2O2 → 2H2O + O2
動物中的過氧化物酶體
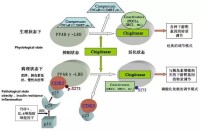
過氧化物酶體
植物中的過氧化物酶體
在植物中過氧化物酶體主要有:①參與光呼吸作用,將光合作用的副產物乙醇酸氧化為乙醛酸和過氧化氫,②在萌發的種子中,進行脂肪的β-氧化,產生乙醯輔酶A,經乙醛酸循環,由異檸檬酸裂解為乙醛酸和琥珀酸,後者離開過氧化物酶體進一步轉變成葡萄糖,這一過程稱為乙醛酸循環,因此植物細胞的過氧化物酶體又稱乙醛酸循環體(glyoxysome)。
過氧化物酶體
從個體發生的角度來看,過氧化物酶體來源於已存在過氧化物酶體的分裂。過氧化物酶體中所有的酶都由核基因編碼,在細胞質基質中合成,在信號肽的引導下,進入過氧化物酶體,引導蛋白質進入過氧化物酶體的信號序列是-Ser-Lys-Leu-COO-。但對於過氧化物酶體膜上與蛋白輸入有關的受體和轉位因子了解甚少,至少和23種被稱為peroxin的蛋白有關,其機理顯著不同於線粒體和葉綠體的蛋白轉運,如受體Pex5(一種peroxin)是伴隨著貨物進入過氧化物酶體的,然後再返回細胞質。
過氧化物酶體病時,血漿、成纖維細胞、羊水細胞中的VLCFA增高。近年來,越來越多的過氧化物酶體病的病種被發現,主要有各型 腎上腺腦白質營養不良(adrenoleukodystrophies),腦肝腎綜合征(Zellweger病),嬰兒型Refsum病,高六氫吡啶羧酸血症(hyperpipecolicacidemia),肢近端型點狀軟骨發育不良(rhizomelic chondrodysplasiapunctata)等。
Zellweger綜合征是一類與過氧化物酶體有關的遺傳病,也叫腦肝腎綜合征,患者細胞的過氧化物酶體中,酶蛋白輸入有關的蛋白質變異,過氧化物酶體是“空的”。腦、肝、腎異常,出生3-6個月內后死亡。

基本信息
- 中文名
- 過氧化物酶體
- 外文名
- peroxisome
- 發現者
- J. Rhodin
- 釋義
- 一種細胞器,存在於一切細胞內
- 時間
- 1954年