共找到2條詞條名為光合碳循環的結果 展開
- 光合碳循環
- 美國卡爾文等人提出的循環過程
光合碳循環
光合碳循環
光合碳循環 photosynthetic carbon cycle 光合作用中碳同化(二氧化碳轉化為糖或其磷酸酯)的基本途徑。又稱卡爾文循環、還原戊糖磷酸循環、還原戊糖磷酸途徑。在綠色植物、藍藻和多種光合細菌中普遍存在。其他碳同化途徑如C4途徑和CAM途徑(如景天科酸代謝)所固定的CO2,最終仍須通過光合碳循環才能被還原成糖。因此它是地球上絕大部分有機物形成的必經途徑。
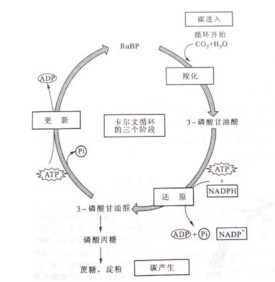
光合碳循環,是一種類似於三羧酸循環(又稱為克雷布斯循環,Krebs cycle,或稱檸檬酸循環)的新陳代謝過程,是光合作用中暗反應的一部分,反應場所為葉綠體內的基質,可使碳以二氧化碳的形態進入發生羧化、還原和二磷酸核酮糖再生共三個階段的循環后,最終以糖的形態離開該循環。整個循環是利用ATP作為能量來源,並以降低能階的方式來消耗NADPH,如此可增加高能電子來製造糖。
M.卡爾文及其同事A.A.本森等從1946年開始,應用新問世的14C標記的14CO2示蹤,並結合紙層析技術,研究了小球藻、柵藻等進行光合作用時碳同化的最初產物,從雙相紙層析放射自顯影的圖譜中看到從14CO2形成了20餘種帶14C標記的化合物。將照光時間漸次縮短至秒級,外推至零時,判斷其中最早出現的是磷酸甘油酸(PGA)。從14C在各同化分子中不同原子間分佈的順序,推斷出C3,C4,C5,C6,C7,糖磷酸酯之間的關係以及它們從PGA形成的先後順序。根據①植物從光轉暗時核酮糖-1,5-二磷酸(RuBP)水平急劇下降,PGA迅速上升,②驟然降低CO2濃度時,PGA迅速下降,RuBP、核酮糖-5-P(Ru5P)、甘油醛-3-P(GAP)依次上升,斷定RuBP是PGA的前體。③找到了催化RuBP與NaH14CO3形成PGA的RuBP羧化酶,肯定RuBP是CO2的受體。CO2同化需要光化學反應形成的ATP及NADPH。整個循環的闡明共花費了9年左右的時間。卡爾文為此獲得了 1961年度的諾貝爾化學獎。1955年E.拉克用菠菜葉片無細胞酶製劑,添加甘油醛-3-脫氫酶,R5P,NAD等,在溶液中用CO2與H2合成了碳水化合物果糖-6-磷酸。
光合碳循環中的十幾個步驟可分為3個部分:
①羧化作用:由RuBP羧化酶催化,將CO2加到RuBP的C-2上,形成中間產物2-羧基-3-酮基核糖醇-1,5-二磷酸,然後水解為兩個分子的3-PGA;
②還原作用:兩個3-PGA經 PGA激酶作用,消耗兩個 ATP,形成兩個1,3-DPGA,再經GAP脫氫酶催化,消耗兩個NADPH,還原為兩個GAP;
③CO2受體RuBP的再生;每 3個RuBP與3個CO2形成6個GAP,5個GAP經過一系列的異構化、縮合與重組,消耗3個ATP,再合成3個RuBP,凈生產一個GAP。
GAP是合成各種有機物質的碳架,可在葉綠體中合成澱粉等物質,又可透過葉綠體被膜上的起跨膜傳遞作用的蛋白稱為P1-運轉器輸出葉綠體外,合成蔗糖等物質。特有的酶氧化戊糖磷酸途徑酶系統的發現促進了光合碳循環各步驟酶系統的分離。其中核酮糖-1,5-二磷酸羧化酶、核酮糖-5-磷酸激酶、景天糖-1,7-二磷酸酯酶是此循環 特有的酶。循環中的RuBP羧化酶、GAP脫氫酶、FBP酯酶、SBP酯酶、Ru5P激酶是調節酶。除一般的代謝調節外,光也起重要的調節作用。光合電子傳遞產生的還原劑使GAP脫氫酶、FBP酯酶、SBP酯酶、Ru5P激酶活化。葉綠體照光時類囊體膜吸收間質中的H引起間質pH值的上升,從pH7.1升至pH8.1;同時類囊體的Mg外流,增加了間質中Mg的濃度,為RuBP羧化酶、FBP酯酶、SBP酯酶、Ru5P激酶催化的反應創造了最適的環境條件。
基本信息
- 中文名
- 光合碳循環
- 所屬領域
- 生物科學
- 作用
- 保證光合作用的穩定進行
- 能量來源
- ATP