肽酶
肽酶
肽酶是一種能夠水解肽鏈的酶,是國際生物化學和分子生物學聯盟命名委員會(Nomenclature Committee of the International Union of Biochemistry and Molecular Biology,NC-IUBMB)推薦的作為所有蛋白水解酶的一般術語,可以分為兩大類型,即內肽酶(E.C.3.4.21~3.4.99)和外肽酶(E.C.3.4.11~3.4.19):其中內肽酶作用於多肽鏈內部的特定肽鍵,如存在於動物消化道的胃蛋白酶、胰蛋白酶和糜蛋白酶等,這些酶對不同氨基酸組成的肽鍵有特異性;外肽酶作用於蛋白質肽鏈兩端肽鍵的酶,作用於氨基端肽鍵的稱氨基肽酶,作用於羧基端肽鍵的稱為羧基肽酶。食物蛋白質在消化道中經過各種肽酶的連續作用,最後分解成為遊離氨基酸。
肽酶通常被俗稱為蛋白水解酶。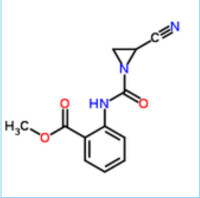 (肽酶與蛋白酶的區別是:蛋白酶將蛋白質水解為多肽,而肽酶將蛋白質水解為氨基酸。)肽酶是一種能夠水解肽鏈的酶。他們是所有生物存活所必需的一種酶,而且在所有蛋白質的編碼中,編碼肽酶的基因佔了2%。在對500個人的肽酶的調查中發現,有14%的的肽酶可以作為藥物的靶點(Southan,2001)。肽酶在許多生物過程中扮演重要的角色,包括消化食物蛋白、胞內蛋白循環、凝血級聯繫統、抗原提呈作用及活化各種蛋白質,包括酶、肽類激素及神經遞質等。
(肽酶與蛋白酶的區別是:蛋白酶將蛋白質水解為多肽,而肽酶將蛋白質水解為氨基酸。)肽酶是一種能夠水解肽鏈的酶。他們是所有生物存活所必需的一種酶,而且在所有蛋白質的編碼中,編碼肽酶的基因佔了2%。在對500個人的肽酶的調查中發現,有14%的的肽酶可以作為藥物的靶點(Southan,2001)。肽酶在許多生物過程中扮演重要的角色,包括消化食物蛋白、胞內蛋白循環、凝血級聯繫統、抗原提呈作用及活化各種蛋白質,包括酶、肽類激素及神經遞質等。
肽酶
許多工業化使用的肽酶都是混合酶,純化的肽酶用量較少。肽酶最早用於乳酪的製作,後來早期用於果汁凝乳(根據荷馬史詩“伊利亞特”中的記載,使用過無花果汁,包含Ficain酶c01.006)和“凝乳酶”,動物胃中含有凝乳酶(a01.006)。肽酶也可用來嫩化肉類,澄清啤酒,提高乳酪和寵物食品的風味。在皮革工業中主要用於要去除的毛髮,使皮革更柔軟(“軟化”和“浸泡”皮革工藝);然而,許多這些專有產品的序列和有機物的來源都不是公開的。肽酶也廣泛使用於清潔材料中,如生物洗滌粉和鏡頭清潔液。除了作為藥物的靶點外,肽酶可以幫助消除胃腸道寄生蟲(Anthelminthics)、幫助燒傷患者去除死皮(清創)、測定血團及通過消化軟骨以緩解背痛、制療椎間盤突出(化學溶核術Chemonucleolysis)等。肽酶在實驗室也被廣泛用作抑制蛋白試劑,並製造多肽所需的蛋白序列。更近期的實驗使用肽酶的抑制特異性處理重組融合蛋白,例如,NIa內肽酶從煙草花葉蝕刻病毒(C04.004)中的病毒多聚蛋白的特定位點:ENLYFQ+G/S。在病毒媒介中特定位點引入帶有能夠編碼特殊蛋白介質(或聚組氨酸及一種蛋白)的核苷酸序列片段從而可以通過加入肽鏈內切酶而簡單的將特定蛋白從混合物中區分出來(Kapust等人,2000年)。肽酶使用的一些例子在表1中列出;Rao的一篇綜述列出了更多的例子(Rao等人,1998年)。
肽酶在生物學、醫學研究和生物技術方面是非常重要的一種酶。因為肽酶活力的調節是一種非常重要的方面,有上百種的蛋白都在作為抑製劑而被同時研究著。
肽酶有多種分類方法,目前普遍使用的是生物化學與分子生物學國際聯盟命名委員會(NC-IUBMB)和由人類基因命名委員會一起推薦的《酶的標準命名法》。某些舊的分類方式因為習慣仍在使用。
根據催化類型可將肽酶大致分成六類:絲氨酸型、蘇氨酸型、半胱氨酸型、天冬氨酸型、谷氨酸型和金屬離子型肽酶。
從生物的角度來說,肽酶還可以消化多肽。
肽酶是能分解蛋白短肽的酶類總稱,大多存在於微生物細胞內,在發酵過程中當細胞自溶后,便釋放肽酶水解肽類形成氨基酸,作為風味前體。他們是所有生物存活所必需的一種酶,而且在所有蛋白質的編碼中,編碼肽酶的基因佔了2%。而按照肽酶的EC系統推薦的作為所有蛋白水解酶的一般術語,可以分為兩大類型,即內肽酶(E.C.3.4.21~3.4.99) 和外肽酶(E.C.3.4.11~3.4.19) ,具體的包括:氨肽酶、二肽酶、二肽基肽酶、肽基二肽酶、絲氨酸型羧肽酶、金屬型羧肽酶、半胱氨酸型羧肽酶、絲氨酸內肽酶、半胱氨酸內肽酶、天冬氨酸內肽酶、金屬內肽酶、蘇氨酸內肽酶、未知類型的內肽酶等。
內肽酶,也就是通常所說的蛋白酶,主要作用於蛋白質多肽鏈內部的肽鍵使蛋白質長鏈分解成短肽片段。蛋白酶根據結構同源性分為不同的宗族,亞類進一步根據序列同源性程度分為不同的家族。
蛋白酶水解蛋白質時,作用部位因肽鍵種類而異。如胰蛋白酶的切點是羧基側為鹼性氨基酸(精氨酸、賴氨酸)肽鍵;胃蛋白酶要求切點兩端有芳香族氨基酸;枯草桿菌鹼性蛋白酶要求切點的羧基側為疏水性芳香族氨基酸(色氨酸、酪氨 酸、苯丙氨酸)。這種蛋白酶對切點的特異性要求叫做蛋白酶的底 物專一性。利用蛋白酶的底物專一性可以定向獲得特殊結構的多肽。
外肽酶廣泛分佈在動物、植物、微生物以及土壤中,在生物正常生理過程和病理生理過程都能起到重要的作用,主要涉及信號轉導、肽類激素的調節、蛋白質結合和消化等過程。研究最多的氨肽酶是乳酸菌氨肽酶,已經從乳酸菌中分離純化出100多種氨肽酶,但其他與食品發酵有關的微生物外肽酶的研究報道相對較少。
外肽酶的分類
酶學委員會 (E. C.)列出了72種不同的外肽酶(http://www.expasy.ch/enzymes)。外肽酶只是在靠近多鏈的一側末端起作用,在遊離N端起作用的酶釋放出單個的氨基酸殘基、二肽或三肽(即分別為氨肽酶、二肽基肽酶和三肽基肽酶);在遊離C末端起作用的酶釋放出單個的氨基酸殘基(羧肽酶)或二肽(二肽基羧肽酶 )。其他外肽酶特異性水解二肽(二肽)或除去被異構肽鍵替換的、環化的,或交聯的末端殘基(即不是α羧基與α氨基的肽鍵)。
外肽酶的命名和性質特點E.C已經建立了從乳酸菌氨肽酶發展起來的微生物氨肽酶系統命名法。對於認識尚未達到基因水平的按照其生化名字或縮寫,其後按發現的順序編號,如LepⅠ、Ⅱ和Ⅲ。如果認識已達到基因水平,就用Pep命名,其後跟上表示特徵的詞的首字母,如Pep N。它是一種金屬氨肽酶,較適於作用N-末端含有脂肪族或帶負電荷的殘基 (Lys,Arg,Leu) ,但是不能作用於含有Pro的肽。谷氨醯胺肽酶和Pep A較適於作用N-末端為Glu和Asp的肽。二肽酶,如Pep D 和Pep V,均表現出對N-末端大的疏水性氨基酸殘基的特異性。
亮氨酸氨肽酶對含有疏水性氨基酸末端殘基的底物具有廣泛特異性。然而,它對N-末端含有精氨酸、賴氨酸或脯氨酸的肽作用很弱。大多數氨肽酶的最適pH在中性範圍,最適溫度範圍37~50℃。植物和動物來源的氨肽酶平均分子量大於100 kDa。氨肽酶的活性會受到Bestain的抑制,抑制的程度隨著蛋白質濃度和處理時間的增加而加強。
絕大多數羧肽酶是金屬羧肽酶,一般都是單個多肽鏈。相對於氨肽酶來說,羧肽酶具有較廣的最適pH範圍(4~8.5)和最適溫度(25~80℃)。與氨肽酶命名相比,羧肽酶的命名還沒有統一的說法,可以根據來源和發現的順序進行命名,如羧肽酶Y(酵母)、羧肽酶W(小麥)等。也有根據酶對底物作用的特性來命名,如Gly-Xaa羧肽酶、絲氨酸型D-Ala-D-Ala羧肽酶等。由於二者都屬於蛋白酶,多數氨肽酶和羧肽酶的活性都會受到EDTA或1,10-鄰菲啰啉的抑制,而合成的賴氨醯丙氨酸比EDTA對含鋅的羧肽酶A抑制作用更強。
外肽酶的結構
氨肽酶的結構
許多氨肽酶是鋅金屬酶類,鋅離子與底物的配基有關。多數微生物氨肽酶是單鏈多肽,其他的含有2、4或6個亞基。一些氨肽酶和羧肽酶含有糖,但都不含脂蛋白。X射線結晶方法揭示了牛晶狀體LAP的三維結構,表明它是一種在活性部位含有2個鋅離子的金屬酶,而哺乳動物間LAP具有結構相似性,不過也發現牛晶狀體LAP和惡臭假單胞菌氨肽酶(ppLAP)之間及其相似,尤其是大腸桿菌和釀酒酵母LAP與牛晶狀體LAP有顯著的氨基酸序列同源性。釀酒酵母氨肽酶1(Ape1)是一種貨物蛋白,在液泡中成熟形式為Ape1 (mApe1),以四面體十二聚合物形式存在。
羧肽酶的結構
對於羧肽酶結構的認識,人們較為熟悉的是CPA和CPB的三維晶體構型。X射線掃描結果表明,牛CPA與CPB的結構十分相似。CPA存在於哺乳動物胰臟中,分子質量34.6 kD,每個酶分子含有1個Zn離子作為輔基,酶蛋白是單一的多肽鏈,約有300個氨基酸殘基。牛CPB分子也含有1個Zn離子,肽鏈有308個氨基酸殘基,其中49%的氨基酸順序與羧肽酶A相同,三維結構也非常相似。與Zn離子配位的氨基酸殘基是His69、Glu72和His196,其催化活性部位包括束縛C-末端羧基的Arg145、作用於肽鍵的Glu270和Tyr248。不同的是,CPB的活性部位還包括Asp255,而CPA的Ile255不是活性部位。CPB的三斜晶體結構顯示,Tyr248朝向溶劑水分子暴露,從而使該酶的疏水性和親水性之間存在細微的平衡,甚至會影響到底物結合以及酶的動力學效率。根據酶的結構差異,金屬羧肽酶分為兩大類:Cowrins和Funnelins類,Cowrins類由原生動物、原核細胞和哺乳動物酶組成,都與溶神經素和血管緊張素轉移酶有關,其催化域殘基數為500~700,參與底物綁定的結構中含有1個由17個螺旋和1個3股的β-摺疊片層。而Funnelins類包括和原型牛羧肽酶A1結構相關的蛋白以及具有特定羧肽酶活性的哺乳動物、昆蟲和微生物蛋白,其催化域內的殘基數在300個以內,而且每邊8個螺旋上側面疊有一個八股β-摺疊片層。Occhipinti等通過定位突變和小角度X射線掃描技術證實了建立分子模型來研究硫化葉菌羧肽酶的三維結構的可行性和條件。
反應特性
所有的外肽酶作用的底物N-末端必須有1個遊離的氨基,或者C-末端必須有1個遊離的羧基,或者兩種結構都需要,而且,外肽酶從肽鏈末端水解1個肽鍵時,釋放的殘基不會多於3個。多肽末端的特定氨基酸、與肽鍵相鄰的氨基酸殘基的性質、多肽鏈的結構和長度共同決定外肽酶的活性。大多數氨肽酶的最適pH值在中性範圍,最適溫度介於37~50℃;與前者相比,羧肽酶具有較寬的作用pH值範圍(4.0~8.5)和較寬的溫度範圍(25~80℃) 。
對食品風味形成的影響
可可
可可豆發酵過程中分泌1種佔優勢的內肽酶(天冬氨酸內肽酶,最適pH3.5)和1種羧肽酶(最適pH5.8)。可可羧肽酶不能水解羧基末端的Arg、Lys 和Pro殘基,較適作用於疏水性氨基酸,對酸性氨基酸水解速度很慢。這2種酶作用生成的疏水性遊離氨基酸和親水性肽,產生可可特有的香味前體物。
牛肉
在肉類生產中,電刺激引起肌肉中外肽酶的活性發生變化,生成的遊離氨基酸種類和含量會影響肉的品質。在牛肉的熟化過程中,對胴體採用低電壓刺激,能夠活化肌肉中的二肽基肽酶的活性,抑制亮氨酸氨肽酶、蛋氨酸氨肽酶和焦谷醯胺氨肽酶的活性,但是對丙氨酸氨肽酶和精氨酸氨肽酶的活性沒有影響。
鹽腌肉
干腌火腿中氨肽酶活性為原來新鮮的火腿的25%~75%,Glu、Leu、Arg、Ala、Tyr 和Lys的含量明顯增加。干腌過程中遊離氨基酸的增加是氨肽酶的作用所致。新鮮豬肉中蛋白酶的組成對干腌火腿產品的品質有很大影響。越苦的樣品中蛋白質分解程度、組織蛋白酶B和二肽醯氨肽酶的活性越強。相反,不太苦的新鮮火腿和腌製成熟的肌肉中氨肽酶活性更高。有苦味的火腿中Met 和Asp含量較高。在腌制加工整個過程中,LAP活性一直都很高。豬骨骼肌中分離純化出的丙氨酸氨肽酶對4-位和5-位上的Ala 有較高的特異性,但缺乏對2-位上的Ala 的特異性。所以,在肉類加工中就會產生二肽和遊離氨基酸,並對肉的最終風味產生影響。促進風味形成的新方法是加入從傳統腌制鹽滷中分離出來的微生物,這些菌產生的脂肪分解酶和蛋白質水解酶與風味的形成有關。
啤酒
麥芽至少含有8種不同的外肽酶,其中有3種羧肽酶,最適pH4.8~5.7;5種氨肽酶,最適pH5.8~8.6。在45℃時肽酶活性穩定。糖化時,溫度從45℃上升到70℃,這些外肽酶迅速被滅活。55℃保溫30min,5種氨肽酶以不同的速度滅活,而羧肽酶活性仍然有70%~100%,其中1種羧肽酶在70℃保溫60min還有40%的殘留酶活。因此,麥芽糖化中,羧肽酶比氨肽酶有更高的熱穩定性。這種特性對糖化時遊離氨基酸的生成極其重要,因為糖化過程形成的遊離氨基酸佔到遊離氨基酸釋放總量的50%~75%。這3種羧肽酶分子質量相似,離子特性和專一性略有差異。麥芽羧肽酶較適作用於含有芳香性氨基酸殘基和Pro的肽。羧肽酶可以降低從胚乳細胞壁提取出來的β-葡聚糖的黏度,並通過水解細胞中的肽來提高胚乳的增溶性。最適pH值和熱穩定性表明羧肽酶不僅可以作用於大麥醇溶蛋白,也可以作用發芽和糖化過程中形成的肽,促使遊離氨基酸的釋放。
乳酪
除了微生物細胞,乳酪提取物中也有胞內外肽酶活性。微生物自溶后釋放出來的外肽酶對酪蛋白寡肽的降解起著很重要的作用。用卡門柏青黴和白地霉製作軟質乳酪時,乳酪的熟化主要受蛋白質水解酶的控制。膠囊處理的重組氨肽酶可以縮短切達乳酪發酵過程,並且能夠提高乳酪的感官評價分值。
對蛋白質水解產物脫苦的影響
從動物來源(奶、肉和魚等)和植物來源(豆類、穀物)的食品蛋白質在食品加工過程中都要發生一定程度的水解。水解作用主要通過兩條途徑:一是細菌或真菌的內肽酶和外肽酶的發酵,二是對蛋白質水解產物的直接加工。苦味肽的存在是大多數蛋白質水解產物的缺陷。蛋白質經內肽酶作用后,形成的水解產物通常都會有苦味,不同的內肽酶由於切點差異,形成的苦肽也各不相同。
商品型外肽酶絕大多數是氨肽酶,主要用於食品蛋白質水解產物的生產過程中,目的在於提高適口性、功能性和營養特性。乳酪生產中通常使用外肽酶降解酪蛋白,並減弱苦味。一般的商業酶最適pH值都在中性範圍。在說明書上酶活都是依據亮氨酸氨肽酶標示,因此,很難比較商用外肽酶的活性。有些酶製劑是外肽酶型,如Protease R、Protease M、Flavourzyme和CorolaseLAP等。如帝國生物技術公司(Imperial Biotechnology)提供2 種用於脫苦的外肽酶混合製劑,Accelase和Debirase。Accelase是Lactococcus lactis的氨肽酶,後者是Lactococcus lactis和A. oryzae的氨肽酶提取物。有些是內肽酶和外肽酶的混合製劑。

基本信息
- 中文名
- 肽酶
- 別名
- 蛋白水解酶
- 外文名
- peptidase
- 術語類別
- 生物術語
- 分類
- 絲氨酸型,蘇氨酸型,半胱氨酸型,天冬氨酸型,谷氨酸型和金屬離子型肽
- 定義
- 肽酶是蛋白水解酶的總稱,是作用於肽鍵的酶。
- 作用
- 將蛋白質分解為遊離氨基酸
- 適用領域
- 生物