綠藻門
藻類植物的一門
綠藻門(拉丁學名:Chlorophyta)藻類植物的1門。主要特徵有:光合作用色素是葉綠素和β胡蘿蔔素及幾種葉黃素;貯藏食物主要是澱粉;在生活史中,產生具有頂端著生的、多為2~4根、等長鞭毛的游泳細胞;有性生殖很普遍,為同配、異配或卵配。藻體有單細胞、群體、絲狀體、葉狀體、管狀多核體等各種類型。本門約8600種,從兩極到赤道,從高山到平地均有分佈。絕大多數種類產於淡水,少數產於海水。
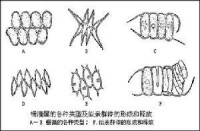
綠藻門
綠藻植物的細胞與高等植物相似,具有真核和葉綠體,葉綠體一至多個,形狀有杯狀、帶狀等(缺葉綠素b時稱載色體),綠藻所含的色素與高等植物相似,也是葉綠素a、葉綠素b以及葉黃素和胡蘿蔔素等,但葉綠素多,因此植物體呈綠色。貯藏的養分為澱粉和油類。澱粉常在葉綠體內的蛋白質(澱粉核)周圍積累,蛋白核有一至多個,它是澱粉形成中心。綠藻的細胞壁成分與高等植物也相似,都是由纖維素構成的。由於綠藻在色素的種類、細胞壁成分、貯藏的養分等方面與高等植物相似,因此多數科學家認為高等植物起源於綠藻。
主要特徵有:①光合作用色素是葉綠素和;和β胡蘿蔔素及幾種葉黃素;②貯藏食物主要是澱粉;③在生活史中,產生具有頂端著生的、多為2~4根、等長鞭毛的游泳細胞;④有性生殖很普遍,為同配、異配或卵配。藻體有單細胞、群體、絲狀體、葉狀體、管狀多核體等各種類型。
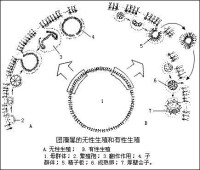
綠藻門
(1)單元單相式:生活史中僅合子階段是二倍體,合子核分裂時即進行減數分裂,然後形成4個游孢子。每一個游孢子發育成一新個體。這種方式在活動的單細胞類型及綠藻綱多數多細胞的物種中比較常見,如衣藻。
(2)單元雙相式:藻體是二倍體,減數分裂發生在形成配子以前,故只有配子是單倍體。雌、雄配子接合成二倍體合子,合子不經減數分裂,直接萌發成二倍體的藻體,如管藻目的種類。
(3)雙元同形:世代交替明顯,由兩個外形相似的不同世代的藻體進行,單倍體的配子體產生配子,配子接合成合子,合子直接發育成二倍體的孢子體。孢子體產生單倍體的游孢子,減數分裂發生在游孢子形成前的第一次分裂,游孢子萌發成單倍體的配子體。如石蓴屬、滸苔屬、剛毛藻屬。
(4)雙元異形:世代交替明顯,也是由單倍體的配子體和二倍體的孢子體交替進行,但這兩個世代的藻體外形不相似。如尾孢藻屬Urospora,多細胞的絲狀體為單倍體的配子體,由它產生配子,配子接合成合子,合子萌發成多核單細胞的二倍體的孢子體。孢子體產生單倍體的游孢子,減數分裂發生在孢子形成時的第一次分裂,游孢子發育成為單倍體的配子體。再如礁膜屬的孢子體是單細胞單核的小藻體,配子體是多細胞的大型藻體。
1.細胞壁
綠藻中除少數物種的細胞原生質裸露、無細胞壁外,絕大多數都有細胞壁。細胞壁由原生質體分泌而成,內層為纖維素,外層為果膠質。果膠為水溶性,外面果膠溶掉再由內面分泌出新的,因此果膠能保持一定的厚度。浮生的藻體因為所分泌的果膠比溶解的多,因此形成膠質套圍於細胞外,這樣就增加了藻體的漂浮能力,有的細胞壁在果膠外還有幾丁質,它可以阻止果膠質的溶解,如剛毛藻屬Cladophora和鞘藻屬Oedogonium;少數物種的內壁不是纖維素而是由胼胝質(callose)組成,如管藻目Siphorales的物種。細胞壁表面平滑,或具顆粒、孔紋、瘤、刺毛等構造,少數物種的細胞壁上還可沉澱石灰質。
2.細胞核
綠藻的大多數物種一個細胞只有1個核,位於細胞中央或邊緣,但是管狀多核體的類型,如松藻屬;或者有些物種在生殖期間出現多核,如傘藻屬Acetabularia在營養時期為單核,進行繁殖時為多核。細胞核有明顯的固定核膜和1個或幾個核仁。在構造上與高等植物沒有什麼不同。核的分裂是有絲分裂和減數分裂,分裂的程序都與高等植物的有絲分裂和減數分裂的程序相似。
3.液泡
液泡都由液泡膜包被,通常較大,位於原生質體中央。但許多團藻目Volvocales、綠球藻目Chloroccales的種類,沒有大的液泡。遊動的藻體內有小而能活動的液泡,在遊動細胞停止活動萌發時,許多小液泡漸漸匯合,最後形成一個大液泡;有些氣生藻沒有大液泡是因為它們生活在缺水的環境內,如原球藻屬Pleurococcus。
4.色素體及色素
色素體是藻類細胞重要的細胞器,形狀多樣。綠藻的色素體有軸生星形、片狀,側生的帶形、網狀、杯狀、粒狀。每個細胞內色素體的數目有1~2個或多個不等。從進化的觀點來看,軸生單個的色素體最原始,側生粒狀色素體最高級。由於後者個體雖小,但接受陽光的相對面積較大,且細胞內色素體數目多,還靠近細胞壁分佈。因此,能接受較多的陽光!光合作用的效能較高。
綠藻色素體所含的色素成分與高等植物相似,有葉綠素、葉綠素b、胡蘿蔔素及葉黃素。但有少數物種所含葉綠素被紅色素(血紅素)遮蔽,因而呈紅色(鹽生杜氏藻Dunaliella salina (Dunal) Teodoresco含有紅色素,含量多時可使細胞呈紅色;多數成熟的合子也含有血紅素,能將葉綠素遮蔽);色素成分的比例也與高等植物相似。光合作用產物也相同,為澱粉。
5.澱粉(蛋白)核
大多數綠藻的色素體內含有1個至數個澱粉核。澱粉核是光合作用產物的儲蓄場所,與澱粉的合成有密切關係。它包含兩部分,中心是蛋白質體,外被澱粉鞘。中心蛋白質體僅是蛋白質的膠黏物,可用染細胞核的染料染色。但也有少數物種並不存在澱粉核,如松藻屬、微孢藻屬Microspora。
6.鞭毛、伸縮泡和眼點
能運動的綠藻和遊動的生殖細胞都具有鞭毛。多數的運動細胞有2根鞭毛,但有些物種有4根或更多的鞭毛。通常游孢子有2根或4根,配子僅有2根,但也有些物種在游孢子的前端有一圈鞭毛,如德氏藻屬Derbesia的物種。
在遊動細胞的前端、鞭毛著生的基部,一般有2個伸縮泡,伸縮泡相互地收縮和展開,收縮時往外排出其中的內含物,擴張時就從原生質體吸收水分而充滿。伸縮泡的功能是排出多餘的水分,同時也排出廢物。不活動的藻體,一般無伸縮泡,但也有少數例外。
綠藻在單細胞和群體的遊動類型物種中基本都有眼點的存在。多細胞物種的游孢子和配子也有眼點。這表示它們的祖先是遊動的。眼點的形狀有環形、卵形或亞線形,一般為橘紅色,生在細胞的前端近鞭毛的基部,也有的生在細胞的中央。眼點結構主要包括一個彎曲的色素板、一個雙面凸的透鏡和在二者之間的一個彎曲無色的感光區。色素板是被假設為具有選擇反射的表面,表面透過長光波(黃、紅色)由它的凹面反射短光波(綠、藍色),其交點集於色素板和透鏡之間的感光層的中間一點。另外,有少數遊動孢子眼點的透鏡在色素層的內面。

綠藻門
1.營養繁殖 群體、絲狀體以細胞分裂來增加細胞的數目。大的群體和絲狀體常由 於動物攝食、流水衝擊等機械作用,使其斷裂。或由於絲狀體中某些細胞形成孢子或配子,在放出配子或孢子后的空細胞處斷裂。或由於絲狀體中細胞間膠質膨脹分離,而形成單個細胞或幾個細胞的短絲狀體(如裂絲藻屬Stichococcus),斷裂的每一小段都可發育成新的藻體。某些單細胞綠藻遇到不良環境時,細胞多次分裂形成膠群體,環境好轉時,每個細胞又可發育成一個新的植物體。
2.無性生殖 無性生殖可形成遊動孢子或靜孢子。遊動孢子無壁,其它構造和衣藻屬的細胞相似。形成遊動孢子的細胞和普通營養細胞沒有區別。有些種的藻體,全體細胞都可產生遊動孢子,但是,群體類型的藻體,不是所有細胞都同時形成遊動孢子。有些藻類僅限於一定的細胞中產生遊動孢子。形成遊動孢子時,細胞內原生質體收縮,形成一個遊動孢子,或經過分裂形成多個遊動孢子,其數目是2的次方。遊動孢子多在夜間形成,黎明時放出,或在環境突變時形成遊動孢子。遊動孢子放出后,遊動一個時期,縮回或脫掉鞭毛,分泌一層壁,成為一個營養細胞,繼而發育為新的植物體。有些藻類以靜孢子進行生殖。靜孢子無鞭毛,不能運動,有細胞壁。另有一種靜孢子,在形態上與母細胞相同,稱似親孢子。在環境條件不良時,細胞原生質體分泌厚壁,圍繞在原生質體的周圍,並與原有的細胞壁癒合,同時細胞內積累大量的營養物質,形成厚壁孢子,環境適宜時,發育成新的個體。
3.有性生殖 有性生殖的生殖細胞叫配子。兩個生殖細胞結合形成合子,合子直接萌發成新個體,或經過減數分裂形成孢子,併發育成新個體。在形狀、結構、大小和運動能力等方面完全相同的兩個配子結合,稱為同配生殖。在形狀和結構上相同,但大小和運動能力不同,大而運動能力遲緩的為雌配子,小而運動能力強的為雄配子,此兩種配子的結合稱為異配生殖。在形狀、大小和結構上都不相同的配子,大而無鞭毛不能運動的為卵,小而有鞭毛能運動的為精子,精卵結合稱為卵式生殖。兩個沒有鞭毛能變形的配子結合,稱為接合生殖。
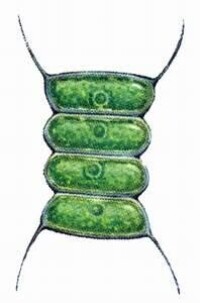
綠藻門
綠藻綱(Chlorophyceae)生活史中具有鞭毛的遊動細胞;有性生殖普遍,但沒有接合生殖。包括12目:團藻目(Volvocales),四孢藻目(Tetrasporales),綠球藻目(Chlorococcales),絲藻目(Ulotrichales),膠毛藻目(Chaetophorales),石蓴目(Ulvales),溪菜目(Prasiolales),鞘藻目(Oedogoniales),剛毛藻目(Cladophorales),
管枝藻目(Siphonocladales),絨枝藻目(Dasycladales)和管藻目(Siphonales)。
接合藻綱(Conjugatophyceae)生活史中不產生有鞭毛的遊動細胞;有性生殖只有接合生殖。此綱只有雙星藻目(Zygnematales)一目。
H.博爾德和M.溫1985年提出把綠藻門分為1綱,16目,即綠藻綱(Chlorophyceae):團藻目(Volvoca-les),四孢藻目(Tetrasporales),綠球藻目(Chlo-rococcales),綠囊藻目(Chlorosarcinales),絲藻目(Ulotrichales),環藻目(Sphaeropleales),膠毛藻目(Chaetophorales),橘色藻目(Trentepohliales),鞘藻目(Oedogoniales),石蓴目(Ulvales),剛毛藻目(Cladophorales),頂管藻目(Acrosiphoniales),雙星藻目(Zygnematales),松藻目(Codiales),管枝藻目(Siphonocladales)和絨枝藻目(Dasyclada-les)。
對綠藻門的進化知之甚少,綠藻的祖先仍不清楚。綠藻門的進化趨勢,根據F.F.布萊克曼1900年的意見,最原始的可能是單細胞種類,由此分出3條進化路線:①自群體的到多細胞的團藻目;②由四孢藻目到絲狀體、扁平葉狀以至杯狀和管狀的各類,高等綠色植物被認為起源於這一分支中的鞘毛藻類(coleochaetes);③綠球藻目這一支失去真正的營養性細胞分裂的種類。已知的綠藻化石不少,尤其是絨枝藻類,最早的記錄是前寒武紀的。
綠藻分佈在淡水和海水中,海產種類約佔10%,淡水產種類約佔90%。有些種類專門生活在海水中,如鞘藻目和雙星藻目。石蓴目和管藻目是海產種佔優勢。絲藻目是淡水種佔優勢,另外也有不少種生活在半鹹水中。海產種多分佈在海洋沿岸,往往附著在10公尺以上淺水中的岩石上。許多海產種有一定的地理分佈,這是由於水的溫度決定的。淡水種的分佈很廣,江河、湖泊、溝渠、積水坑中,潮濕的土壤表面,牆壁上,岩石上,樹榦上,花盆四周,甚至在冰雪上都可找到。它們中部分是沉在水中生活,許多單細胞和群體種類是漂浮在水中,但在海水中沒有浮遊的綠藻,有的綠藻也可以寄生在動物體內,或者與真菌共生形成地衣。一般淡水種不受水溫的限制,大部分分佈在世界各地。
綠藻是藻類植物中最大的一門,約有350個屬,5000—8000種。分成兩個綱,即綠藻綱和輪藻綱。我國一般將綠藻綱分為13個目,即團藻目、四孢藻目、綠球藻目、絲藻目、具毛藻目、石蓴目、溪菜目、鞘藻目、剛毛藻目、管藻目、管枝藻目、絨枝藻目和接合藻目。輪藻綱中只有輪藻目。
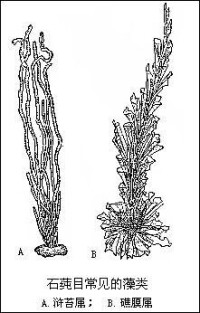
綠藻門
(1)衣藻屬(Chlamydomonas)是團藻目內單細胞類型中的常見植物。本屬約有100種以上,生活於含有機質的淡水溝和池塘中,早春和晚秋較多,常形成大片群落,使水變成綠色。
植物體為單細胞,卵形,細胞內有1個厚底杯狀的葉綠體,其底部有1澱粉核。細胞核位於葉綠體上方的杯中。藻體的前端有2條等長的鞭毛,其基部有2個伸縮泡,旁邊有1個紅色眼點。在電子顯微鏡還可以看到類囊體、線粒體和高爾基體等。
衣藻通常進行無性生殖。生殖時藻體常靜止,鞭毛收縮或脫落,變成遊動孢子囊。原生質體分裂為2、4、8、16,各形成具有細胞壁和2條鞭毛的遊動孢子(zoospore),囊破裂后,遊動孢子逸出發育成新個體。
衣藻的有性生殖多數為同配生殖。原生質體分裂成8~64個小細胞,稱配子(gamete)。配子在形態上和遊動孢子相似,只是體形較小。配子從母細胞中放出后,遊動不久即成對結合,成為2N、具4條鞭毛的合子,合子遊動數小時后變圓,形成有厚壁的合子。合子經過休眠,在環境適宜時萌發。萌發時經過減數分裂,產生4個遊動孢子。當合子壁破裂后,遊動孢子游散出來各形成一個新的衣藻個體。
(2)團藻屬(Volvox)屬於團藻目。春夏兩季常見生於淤積的淺水池沼中。植物體是由數百至上萬個衣藻型細胞組成的球形群體,衣藻型細胞排列在球體的表面,空心球體內充滿膠質和水。有的種有胞間連絲,逐步過渡到多細胞的個體。群體中只有少數大型的細胞能進行繁殖,稱此為生殖胞(gonidium)。無性生殖時,少數大型的生殖胞經多次分裂形成皿狀體(plakea),再經翻轉作用(inversion)發育成子群體,落入母群體腔內,母群體破裂時放齣子群體,即為一新植物。有性生殖為卵式生殖,精子囊和卵囊分別產生精子和卵,精子和卵結合形成厚壁的合子。當母體死亡腐爛后,合子落入水中,休眠后經減數分裂,發育成一個具有雙鞭毛的遊動孢子,逸出后萌發成一新的植物體。
小球藻和珊藻富含蛋白質可供人食用和作動物飼料。綠藻是藻類生理生化研究的材料及宇宙航行的供氧體,有的可制藻膠。綠藻在水體自凈中起凈化和指示生物的作用。

基本信息
- 中文學名
- 綠藻門
- 拉丁學名
- Chlorophyta
- 界
- 植物界
- 門
- 綠藻門
- 分布區域
- 淡水中為最多,流水和靜水中都可見到;陸地上的陰濕處和海水