外顯子
外顯子
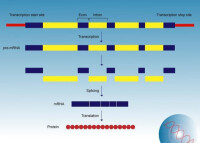
斷裂基因中的編碼序列。外顯子(expressed region)是真核生物基因的一部分,它在剪接(Splicing)后仍會被保存下來,並可在蛋白質生物合成過程中被表達為蛋白質。外顯子是最後出現在成熟RNA中的基因序列,又稱表達序列。
既存在於最初的轉錄產物中,也存在於成熟的RNA分子中的核苷酸序列。術語外顯子也指編碼相應RNA外顯子的DNA中的區域。所有的外顯子一同組成了遺傳信息,該信息會體現在蛋白質上。
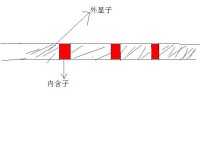
外顯子和內含子
在反式剪接中,不同mRNA的外顯子可以被接合在一起。外顯子在剪接(Splicing)后仍會被保存下來,並可在蛋白質生物合成過程中被表達為蛋白質。簡言之,外顯子就是指真核細胞的基因在表達過程中能編碼蛋白質的核苷酸序列。關鍵概念:比較不同物種的相關基因,發現相應的外顯子序列通常是保守的,而內含子序列則很少保守。編碼蛋白質的序列通常處於選擇壓力之下,內含子由於沒有選擇壓力,因此比外顯子的進化快得多。通過確定在多種生物中出現的片段來鑒定編碼區域,而外顯子的保守性可以作為這種鑒定的基礎。
人類大部分基因組序列都是被垃圾DNA序列分隔成一段段,給定一個已知的目標蛋白質和基因組序列,在該基因組序列中找出一組子字元串(候選外顯子),使得其拼接(剪接)與目標蛋白質最匹配(即去掉垃圾DNA序列)。一個強力方法是尋找基因組序列與目標蛋白質序列間的所有局部相似性。若第一個取自基因組序列的子字元串展示了充分相似性於目標蛋白質,那麼這個子字元串可被認為是一個推定的外顯子。將推定外顯子結構化為基因組序列中的賦權區間,它可用三個參數(l、r、w)來描述,l、r分別是推定的外顯子的左邊、右邊的位置,w為其權重。權重w可反該區間是一個外顯子的可能性。鏈是不重疊賦權區間的任一集合,一個鏈的總權重是該鏈中所有區間的權重之和。給定一個推定的外顯子集,尋找非重疊的推定的外顯子的一個最大集。輸入:賦權區間(推定的外顯子)集。輸出:該集合中區間的最大。

基因啟動子區域以及外顯子-內含子連接處cfDNA降解特徵
操作步驟及其基本原理是:
⑴基因組DNA經“霰彈法”切成小片段后,克隆在位於“外顯子捕捉序列”下游的克隆位點上。
⑵將這些重組載體匯總後感染反轉錄病毒的專宿包裝細胞系(ecotropicretroviralpackagingcellline)——ψ2細胞系。ψ2細胞提供蛋白質產物使載體(自身不能合成病毒蛋白質)成為反轉錄病毒在細胞里增殖。當反轉錄病毒在細胞內轉錄時,如果插入片段中包含有功能的SA位點,則有可能發生RNA剪接反應而將ⅣS切除。
⑶已剪接和未剪接的病毒RNA都包裝在病毒子(virion)中,從細胞培養液中收集後用來感染兼宿反轉錄病毒包裝細胞系(amphotropicretroviralpackagingcellline)PA-317。這使反轉錄病毒再進行一輪複製,併產生能感染猴腎細胞系COS細胞的高效價病毒原種。這樣做是由於上一輪克隆在病毒中的插入片段的剪接效率極低,而在第二輪複製時則大大提高了RNA剪接的機會。
⑷從第二個細胞系PA-317細胞中分離得到的病毒,用來感染組成型產生SV40T(腫瘤)抗原的COS細胞。病毒RNA基因組被反轉錄,並在載體上的SV40複製起點作用下,以環狀DNA附加體形式進行複製。
⑸從COS細胞中回收複製的附加體DNA,經限制性內切酶DpnI酶切後轉化細菌。在含卡那黴素(Kn)和5—氯—4—溴—3—吲哚—β—D—半乳糖苷(X-gal)的培養基上挑選轉化子。盧—半乳糖苷酶可水解X—gal而生成藍色產物。因此,不產生β—半乳糖苷酶的轉化子菌落則呈白色。
⑹只挑選出白色菌落作進一步研究的材料。白色菌落的生成可以有二種原因。一是由於基因發生突變,使夕—半乳糖苷酶失去活性;二是由於在反轉錄病毒生活周期的RNA時期中發生了剪接反應,從而丟失了α,β—半乳糖苷酶基因。
⑺如果是基因突變,則大多數將是缺失了載體中的“外顯子捕捉”部分,就可用人的口—珠蛋白基因片段為探針作菌落雜交,很快可得到驗證。
⑻如果是真正發生了RNA剪接事件,準確的剪接反應可切除作為標記的ⅣS,使人口—珠蛋白基因的第1外顯子與落入了捕捉陷阱的插入片段中的外顯子序列連接,這可直接測定其序列加以證明。從捕捉到的外顯子出發,就可進一步用作探針去從基因組基因文庫或cDNA文庫中分離出基因。
應用聚合酶鏈反應-單鏈構象多態性(polymerasechainreaction-singlestrandconformationpolymorphsim,PCR-SSCP)及DNA直接測序技術檢測68例SAD患者和65名正常老年人的早老素-1基因第5外顯子。
外顯子圖片
多數早發性家族性阿爾茨海默氏病(familialalzheimer'sdisease,FAD)與14號染色體上的早老素-1(presenilin-1,PS-1)基因突變有關,少數早發性FAD與PS-2基因突變有關。早老素基因與散發性Alzheimer病(sporadicalzheimer'sdisease,SAD)的關係,目前國內外研究較少,為探討PS-1基因突變在SAD發病機理中的作用,我們用聚合酶鏈反應-單鏈構象多態性(polymerasechainreaction-singlestrandconformationpolymorphism,PCR-SSCP)、DNA直接測序技術檢測了68例SAD患者及65名正常老年人PS-1基因第5外顯子,發現在SAD患者中也存在PS-1基因第5外顯子突。
外顯子圖片
AdrianKrainerandLucaCartegni為了在外顯子跳躍突變中恢復正常的外顯子,將一個合成的RS(精氨酸-絲氨酸)結構域融合到能夠與特異性外顯子結合的反義核苷酸片段上。他們首先檢測了ESSENCE方法對乳腺癌基因1(BRCA1)突變的作用,BRCA1在外顯子18上的ESE突變會導致外顯子跳躍。他們發現將ESSENCE化合物與能夠與外顯子18反義結合的片段融合作用后在體外能夠恢復正常的剪切,其中反義片段和RS片段都是必需的。
研究者接著對另一種外顯子跳躍突變導致的疾病模型進行研究。神經退行性疾病脊髓肌肉萎縮由運動神經元存活基因1(SMN1)的功能性拷貝缺失導致。SMN1的缺失可以有SMN2的功能所補償,但SMN2的外顯子7的單核苷酸突變會導致整個外顯子缺失。研究者發現用ESSENCE方法也能在體外恢復SMN2突變缺失的外顯子7。這個新方法雖然還需要體內的許多研究數據,但顯然對相關遺傳疾病的治療確實帶來的曙光。
許多基因中遺傳上的“無義”片段--即內含子,會妨礙基因指導蛋白質的合成。現在,一篇發表於3月11日期的《自然遺傳學》雜誌上的文章提出了基因識別這些內含子的新機制。
外顯子圖片
為進一步驗證實驗結果,Baralle的研究小組將含有A-T病人中缺失的這個鹼基對,共由12鹼基對組成的健康基因序列插入到一個完全不同基因的外顯子中。令人驚異的是,剪接機制開始把整個外顯子當作內含子處理,將該外顯子剪切掉。Baralle得出結論:實驗結果表明,這個序列行使作用時,能夠幫助剪接酶識別內含子;缺失時(如在A-T病人中),就會錯移到外顯子中去。Baralle猜測,這個序列可能也控制著其它基因的剪接,對引起包括癌症在內的各種疾病有一定作用。
這項工作標誌著一次“重大發現”,紐約城哥倫比亞大學的JimManley評價道。Manley認為內含子中間的這段關鍵序列也許是幫助剪接機制識別和除去內含子的“踏腳石”。在整個基因組中,可能存在“幾十個類似機制”來確定內含子的組成
基本信息
- 中文名
- 外顯子
- 外文名
- expressed region
- 基因類別
- 真核生物基因
- 術語含義
- 編碼相應RNA外顯子的DNA中的區域