瑪君龍
阿貝力龍科獸腳類恐龍
瑪君龍(屬名:Majungasaurus)又譯瑪宗格龍,意為“馬達加斯加的蜥蜴”,是獸腳亞目阿貝力龍科的一屬,生存於白堊紀末的馬達加斯加,約7000萬年前到6500萬年前。目前僅有一個已確認種M.crenatissimus,近期發現瑪君龍最大個體為10米長,4噸,但由於化石太碎,只是疑似瑪君龍,還不能確定。瑪君龍一度被認為是種厚頭龍下目恐龍,名為瑪君顱龍/瑪宗格厚頭龍(Majungatholus),瑪君顱龍現被視為瑪君龍的次異名。

瑪君龍
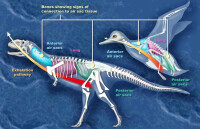
瑪君龍是種中等體型的獸腳亞目恐龍,平均體長為7米左右,包含尾巴在內。大型瑪君龍個體的破碎化石顯示,有一些成年大個體體身長可達8米。科學家們估計成年的瑪君龍平均體重達1500公斤。科學家們估計較大的瑪君龍的體重可以接近其近親食肉牛龍的大小,達到2000公斤。
與大部分的獸腳亞目恐龍相比,瑪君龍的頭顱骨相當著名,而且類似其他阿貝力龍科恐龍的頭顱。如同其他阿貝力龍科的頭顱,瑪君龍的頭顱長度與高度比例較短,但沒有食肉牛龍那樣短。大型成年個體的頭顱骨長度約60到70厘米。前上頜骨使得口鼻部前端非常鈍,這是阿貝力龍科恐龍的典型特徵。然而,與其他阿貝力龍科恐龍的頭顱骨相比,瑪君龍的頭顱骨寬度較寬。所有阿貝力龍科恐龍的頭顱骨都有粗糙不平的表面,瑪君龍也不例外。瑪君龍的鼻骨特別厚,並且互相固定,鼻骨的下半部有個低鼻脊。頭頂的固定額骨有個明顯的半球形角狀物。在瑪君龍存活的時候,這些結構可能由某種結構所覆蓋,可能由角質所構成。電腦斷層掃描顯示這些鼻部結構與額角擁有中空空間,可能用來減輕重量。瑪君龍的牙齒有短齒冠,這是典型的阿貝力龍科特徵;而瑪君龍的上頜骨與齒骨分別有17顆牙齒,是阿貝力龍科當中最多的,除了皺褶龍以外。
瑪君龍的顱后骨骼極度類似食肉牛龍與奧卡龍的顱后骨骼,這兩個屬是其他曾發現完整骨骸的阿貝力龍科恐龍。瑪君龍是二足恐龍,擁有長尾巴,以平衡頭部與胸部,使得重心位在臀部。雖然頸椎擁有許多空間以減輕這些部位的重量,但還是有加大肌肉附著在上面,而肋骨互相連錯、加強,所以頸部還是很結實。頸部肋骨有骨化肌腱附著,如同食肉牛龍。不同的是,瑪君龍的頸部肋骨側邊有長凹處,以漸輕重量。綜合以上特徵,瑪君龍的頸部強壯、充滿肌肉。
肱骨是前肢唯一被敘述的部份,肱骨短而彎曲,極度類似食肉牛龍與奧卡龍的肱骨;這顯示瑪君龍可能擁有非常類似的短前肢與四指。如同其他阿貝力龍科恐龍,瑪君龍的前肢使身體相比,短而結實。瑪君龍的脛骨比食肉牛龍的脛骨還要結實,膝蓋部位有明顯的突出。距骨與跟骨互相固定著,腳部有3根具有功能的腳趾,而最小的第一趾並未接觸到地面上。
瑪君龍有一個特殊的頭顱骨,它們的頭顱骨又長又厚,表面粗糙不平。瑪君龍的口鼻部很短,前肢短小,後肢長而粗壯。瑪君龍的頭頂上有一個半球形角狀物,這種角狀物可能由角質構成,並由某種結構覆蓋 。
瑪君龍目前已發現數個保存良好的頭顱骨與大量的完整骨骸,並在近期成為南半球的獸腳亞目恐龍中最廣受研究的一種。瑪君龍似乎與印度的阿貝力龍科恐龍關係較近,而離南美洲與非洲的阿貝力龍科恐龍較遠,這點在生物地理學上有重要的意義。瑪君龍是該生態系統的頂級掠食者,主要獵食蜥腳下目恐龍,例如掠食龍;瑪君龍也目前唯一有直接證據顯示同類相食現象的恐龍。
瑪君龍被分類於獸腳亞目的阿貝力龍科演化支,而阿貝力龍科與西北阿根廷龍科共同組成阿貝力龍超科,阿貝力龍超科則是角鼻龍下目的一個分支。阿貝力龍科恐龍的特徵是:高頭顱骨、鈍口鼻部、面部骨頭的凹凸不平表面(與鯊齒龍科的趨同演化結果)、縮短的前肢(與暴龍科的趨同演化結果)、以及結實的後肢。
如同許多的恐龍科,阿貝力龍科內部成員的演化關係非常混淆。數個親緣分支分類法研究顯示在系統發生學上,瑪君龍與南美洲的食肉牛龍有接近親緣關係,而其他屬則無法確定演化位置。一個最近的研究,使用最完整的資訊,不將瑪君龍與印度的勝王龍、印度龍分類於同一演化支,而是與南美洲的食肉牛龍、肌肉龍、爆誕龍、奧卡龍、阿貝力龍、以及非洲的皺褶龍分類於同一演化支。這個研究使得阿貝力龍科可能別在西岡瓦納大陸與東岡瓦納大陸形成個別的次演化支。需要更進一步的發現與研究才能協助了解阿貝力龍科與瑪君龍的詳細系統發生學。
雖然瑪君龍沒有什麼特殊的影視作品但瑪君龍首次出現於紀錄片(侏羅紀格鬥場)在影片中被描述成同類相殘的肉食恐龍,同樣還詳細的講解了瑪君龍的交配和生活。瑪君龍也出現在2011紀錄片(恐龍星球)中在影片里描述了瑪君龍龍的弱點和武器,比如它弱小的手臂類似霸王龍。
瑪君龍同樣也出現在手游(侏羅紀世界)中,但被錯誤的描述成2個手。遊戲中瑪君龍可以和鳳凰翼龍合體成為(Hybrid)混種鳳凰瑪君龍。

瑪君龍
法國古生物學家Charles Depéret在1896年敘述了第一個來自於馬達加斯加西北部的獸腳亞目化石。這些化石是由一個法國陸軍軍官在貝齊布卡河(Betsiboka River)沿岸發現,包含兩顆牙齒、一個指爪、以及一些脊椎骨,目前存放在里昂第一大學。Depéret將這些化石分類於斑龍屬的M.crenatissimus種,斑龍屬在當時是個“未分類物種集中地”,包含許多大型獸腳類,但這些物種彼此之間並沒有關聯。M.crenatissimus的種名衍化自拉丁語,crenatus意為“凹槽”,而issimus意為“最”,意指它們牙齒前後的大量鋸齒狀邊緣。Depéret稍後將它們重新歸類於北美洲的傷龍,這是另一個所知有限的屬。

瑪君龍瑪君龍的模式標本
在1993年,紐約州立大學斯通尼布魯克分校與馬達加斯加的安塔納那利佛大學展開了馬哈贊加盆地計劃,挖掘並研究馬哈贊加省貝立佛查村附近的晚白堊紀地層中發現的化石與地質。第一次的挖掘活動發現了數百顆獸腳類恐龍的牙齒,經鑒定后屬於瑪君龍,有些牙齒附著在一個前上頜骨之上,該前上頜骨在1996年被敘述。而接下來的七次挖掘活動發現了數萬件的化石,其中許多化石屬於新的物種。馬哈贊加盆地計劃在該地區發現了原有已知化石物種的五倍。
在1996年,這個挖掘團隊發現了一個保存極良好的完整獸腳類頭顱骨(菲爾德自然歷史博物館PR2100標本)。頭顱的頂部有個半球形隆起物,類似蘇伊士與塔丘特所敘述的瑪君顱龍。在1998年,瑪君顱龍再度被敘述為阿貝力龍科恐龍,而非厚頭龍類。雖然瑪君龍(Majungasaurus crenatissimus)的命名比瑪君顱龍(Majungatholus atopus)還早,但敘述者認為瑪君龍模式標本的齒骨過於零碎,而沒有足夠的把握將它與頭顱骨分類於同一物種。
在接下來10年,發現了一系列較不完整的頭顱骨,以及數十個不同個體的部份骨骸,這些骨骸有幼年體也有成年體。該計劃的成員也發現了數百件瑪君龍的相關骨頭,以及數千顆零散的牙齒。將這些化石組合過後,幾乎拼湊出完整的骨骸,但大部分前肢、大部分的骨盆、以及尾巴的末端等部份仍未發現。這個挖掘計劃在2007年達到頂峰,一份由7個關於瑪君龍的科學研究,所集合的專題論文,在古脊椎動物化石學會的學會會議上發表。這個專題論文是以英文寫成,但每份研究都有以馬拉加斯語寫成的概要。在論文之中,Lavocat將之前提到的齒骨重新鑒定,並將它分類於瑪君龍。因此,瑪君顱龍被瑪君龍所取代。雖然該份專題論文內容非常廣泛,但該論文只敘述從1993年到2001年期間所發現的化石。而在2003年與2005年,在該地又發現了相當數量的標本,有些標本非常完整,這些化石正等待者更進一步的敘述。
瑪君龍最特殊的地方在於頭顱上的裝飾物,包括突出、固定的鼻骨與額角。其他的角鼻龍類,包含食肉牛龍、勝王龍、以及角鼻龍的裝飾物都在頭部之上。這些結構可能作為物種內競爭使用,但正確的功能仍未確定。瑪君龍額角的內部中空空間可能會讓額角脆弱,並妨礙額角使用於直接的撞擊上,所以這些角狀物可能作為展示物使用。但瑪君龍的角狀物在不同個體間有很大變化性,所以沒有證據顯示它們是兩性異形。
對於瑪君龍來說,它們最血腥的事就是同類相食。瑪君龍同類相食的證據來自於它們骨骼化石上的瑪君龍齒痕,這種齒痕與瑪君龍的牙齒大小和間隙完全匹配。瑪君龍身長約20英尺(約合6米),曾經於7000萬年前生活於馬達加斯加平原之上。研究人員介紹說,在當時也存在身體瘦小的靠採摘植物為食的同類。研究數據顯示,瑪君龍不僅僅以靠採摘植物為食的同類為食,有時也以自己的家族同類為食。相對於其他動物來說,恐龍之間的同類相食相對常見,但證據很少。
科學家們提出瑪君龍與其他阿貝力龍科恐龍的獨特頭顱形狀,顯示出與其它獸腳類恐龍不同掠食習性。大多數獸腳類恐龍有長、低矮、狹窄的頭顱骨,阿貝力龍科恐龍的頭顱骨較高且寬,長度通常較短。其他獸腳類恐龍的狹窄頭顱骨可承受強壯咬合時的垂直壓力,但不適合承受扭轉力量。與現代哺乳類掠食者相比,大多數獸腳類恐龍可能使用類似犬科動物的方式獵食,使用長而狹窄的口鼻部多次咬傷獵物,使獵物變虛弱。
阿貝力龍科恐龍,尤其是瑪君龍,可能改採用類似現代貓科動物的方式獵食,使用短而寬廣的口鼻部咬住獵物緊緊不放,直到它們被制服。瑪君龍的口鼻部較其他阿貝力龍科恐龍還寬廣,而且它們的其他生理特徵也支持者這個獵食方式假設。瑪君龍的頸部較壯,且擁有結實的頸椎、交錯的頸部肋骨、骨化的肌腱、以及附著在脊椎骨與頭顱後方的強化肌肉。這些強化肌肉可使瑪君龍的獵物掙扎時,瑪君龍仍能保持頭部的穩定性。阿貝力龍科恐龍的頭顱骨有許多區域,有皮膚經過礦化作用而成的骨頭,形成凹凸不平的表面。阿貝力龍科的鼻骨加厚且固定住,可增加強度,這種特徵在瑪君龍身上特別明顯。另一方面,瑪君龍的下頜兩側各有一個大型洞孔,這特徵可見於其他角鼻龍類恐龍;特定關節間有滑液關節,可允許下頜的大幅度變形擴張,但不能達到蛇類的擴張程度。這些下頜特徵可防止瑪君龍嘴中的獵物掙扎時,下頜遭到破裂。下頜的前部牙齒比其他牙齒還要結實,可提供咬合時的支撐點;而瑪君龍的低齒冠牙齒可防止獵物掙扎時,將牙齒搖斷。最後,不像異特龍與大部分獸腳類恐龍,它們的牙齒呈前後側彎曲;而阿貝力龍科恐龍的牙齒,例如瑪君龍,它們的牙齒前側彎曲,而後側較直。這種牙齒結構可用來將獵物切碎,而非將獵物托在咬合的地方。
瑪君龍是它們所處環境的最大型掠食者,而該地區同一時期的最大型草食性動物是蜥腳類恐龍,例如掠食龍。科學家們認為瑪君龍,或者其他的阿貝力龍科恐龍,特別以蜥腳類恐龍作為獵食對象。它們頭部與頸部的增強,以適應咬合與咬住的攻擊方式,可能對於巨大、強壯的蜥腳類恐龍特別有效。相較於其他獸腳類恐龍較修長的後肢,瑪君龍短而結實的後肢,可支持這個假設。雖然瑪君龍的奔跑速度無法趕上相同大小的獸腳類恐龍時,它們仍可以追上移動緩慢的蜥腳類恐龍。瑪君龍的結實後肢骨頭顯示它們擁有非常強壯的後腿,而後腿長度較短,可降低瑪君龍的重心位置。因此瑪君龍犧牲速度,以增加力量。在掠食龍骨頭上發現的瑪君龍牙齒痕迹,確認它們至少以這些蜥腳類恐龍為食,但不確定瑪君龍是獵食它們,或是在它們死後吞食它們的屍體。
雖然蜥腳類恐龍可能是瑪君龍的食物來源之一,最近在馬達加斯加的發現顯示瑪君龍有同類相食現象。在許多瑪君龍骨頭上發現了牙齒痕迹,類似同一地區蜥腳類化石上的牙齒痕迹。這些牙齒痕迹的間隔與瑪君龍的牙齒間隔相同,牙齒大小也相符合,並擁有較小的凹口,也與瑪君龍的牙齒邊緣符合。因為瑪君龍是該地區已知的唯一大型獸腳類恐龍,最簡易的解釋是瑪君龍有同類相食現象。三疊紀的腔骨龍曾被認為有同類相食現象,但已被否決,使得瑪君龍是唯一確認有同類相食傾向的獸腳類恐龍,但也有證據顯示其他種也有同類相食現象。

化石
瑪君龍與現代鳥類的呼吸系統比較圖
瑪君龍氣囊的發現,除了了解它們的呼吸系統以外,也有助於理解生物的演化。瑪君龍屬於角鼻龍下目,而鳥類屬於堅尾龍類演化支,角鼻龍下目與堅尾龍類在獸腳類演化歷史的非常早期就已經分開演化。而在兩個支系都發現了鳥類的呼吸系統,顯示這呼吸系統在兩個支系分化前就已經存在,而非鳥類獨自演化出來。這個發現更提供了鳥類起源於恐龍的進一步證據。
對於瑪君龍完整頭顱骨(編號FMNH PR2100)的電腦斷層掃描,顯示出大略的腦部與內耳結構。與身體相比,瑪君龍的腦部非常小,除了這方面之外則類似非虛骨龍類的獸腳類恐龍,有非常傳統的外形,較類似現代鱷魚,而非鳥類。瑪君龍與其他獸腳類恐龍的差別之一,是它們的較小型絨球,絨球是小腦的一個區域,可協助調節眼睛與頭部的動作。這顯示瑪君龍與其他擁有小型絨球的阿貝力龍科恐龍,例如印度龍,並非依靠快速的頭部運動與視力來捕抓獵物。
對於內耳的研究,可以推論出瑪君龍的行為模式。內耳的半規管可協助保持平衡,而當頭部保持警戒姿勢時,側半規管會與地面保持平行。瑪君龍的頭顱骨幾乎與地面平行,當它們旋轉頭部時,它們的側半規管會與地面保持平行。當其他獸角類恐龍保持警戒姿勢時,它們的頭部會大幅朝下。與它們的更基礎近親角鼻龍相比,瑪君龍的側半規管明顯地較長,顯示它們對於頭部側向運動,有更強烈的知覺。
一個2007年的研究敘述了瑪君龍骨頭上的病理。科學家們檢驗了至少21個標本,並發現四個明顯病狀。當其他大型堅尾龍類恐龍,例如異特龍科與暴龍科,已經有病狀方面的研究時,瑪君龍是第一個被研究病狀的阿貝力龍超科恐龍。瑪君龍的頭顱骨上沒有發現任何傷口,而暴龍科的面部常出現傷口。其中一個瑪君龍標本的足部掌骨,發現了明顯的骨折與痊癒痕迹。
瑪君龍的大部分病狀位在脊椎上。例如一個幼年個體的一個背椎下側有外生骨瘤。這個骨瘤可能導因於軟骨內骨化或韌帶硬化成骨頭,但不確定骨化的原因。維生素A過高症(Hypervitaminosis A)與骨刺被排除於原因之外,骨瘤也被視為不太可能。在另一個標本的尾椎神經棘上也發現了一個贅生物;神經棘是從脊椎突出來,並提供肌肉附著點。類似的神經棘贅生物也在異特龍與惡龍的身上發現,這些贅生物可能導因於棘間韌帶(Interspinal ligament)或棘上韌帶(Supraspinal ligament)的骨化。
目前所發現瑪君龍的最嚴重病狀位在五個連續的大型尾椎上。前兩個尾椎有少部份變形,骨頭左側有一個大型凹溝。后三個尾椎完全癒合,形成一個大型堅硬骨塊。沒有跡象顯示這些連續尾椎後面還有其他骨頭。科學家們從最後一個尾椎的大小判斷,認為還有約10個尾椎遺失了。一個解釋認為這些病狀是因為尾巴末端遺失而引起的身體創傷,並使剩下的尾椎產生骨髓炎。另一個說法是尾椎先開始感染,並導致尾巴末端壞死,然後脫落。這個病例是目前已知第一個有尾巴截斷截斷現象的非鳥類的獸腳亞目恐龍。

模型骨架
瑪君龍的所有標本被發現於馬達加斯加西北部馬哈贊加省的梅法拉諾組。其中大部分標本,尤其是最完整的一群化石,都發現於Anembalemba段,而下層的Masorobe段與上層的段也發現了瑪君龍的牙齒。這些沉積層並沒有經過放射性定年法檢驗過,但生物地層學與古地磁學顯示該地層年代為馬斯特里赫特階,約7000萬年前到6500萬年前。瑪君龍的牙齒在馬斯特里赫特階結束時被發現,在白堊紀-第三紀大滅絕事件發生時。
在白堊紀末期,馬達加斯加是座島嶼,在2000萬年之前與印度次大陸分離出來。馬達加斯加往北方漂移,但緯度比現代還往南10到15度。白堊紀末期的普遍氣候為半乾旱,在氣溫與降雨量上有明顯的季節變化。瑪君龍棲息於岸邊的泛濫平原,該泛濫平原被許多含砂量高的河道所分隔。強力的地理證據顯示,在雨季的開始時有周期性的土石流流經這些河道,將乾季死亡的生物屍體掩埋,提供了化石化的環境。該地區的海平面在馬斯特里赫特階時期上升,可能持續到古新世,所以瑪君龍可能漫遊於海岸環境,例如潮坪。而鄰近的貝立佛查組則代表了同時代的海相環境。
除了瑪君龍以外,梅法拉諾組所發現的化石還包含:魚類、青蛙、蜥蜴、蛇、7種鱷形超目動物、5或6種哺乳類、早期鳥類Vorona、與數種其他鳥類、可能擁有飛行能力的馳龍科脅空鳥龍、西北阿根廷龍科的惡龍、以及兩種泰坦巨龍類,例如掠食龍。瑪君龍是該地區白堊紀後期的最大型肉食性動物,也可能是優勢掠食者,但大型鱷形超目動物,例如馬任加鱷與Trematochampsa可能會跟瑪君龍競爭水邊的食物。

瑪君龍
