高爾基體
單位膜構成的扁平囊疊加在一起組成的物質
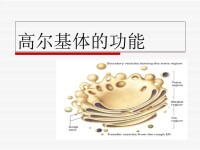
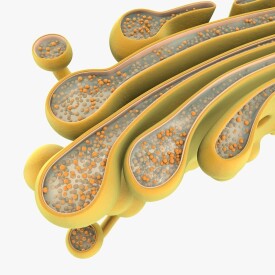
高爾基體是由單位膜構成的扁平囊疊加在一起所組成。扁平囊為圓形,邊緣膨大且具穿孔。一個細胞內的全部高爾基體,總稱為高爾基器。一個高爾基體常具5——8個囊,囊內有液狀內含物。
高爾基體(Golgi apparatus,Golgi complex)亦稱高爾基複合體、高爾基器。是真核細胞中內膜系統的組成之一。為義大利細胞學家卡米洛·高爾基於1898年首次用硝酸銀染色的方法在神經細胞中發現。是由光面膜組成的囊泡系統,它由扁平膜囊(saccules)、大囊泡(vacuoles)、小囊泡(vesicles)三個基本成分組成。
高爾基體
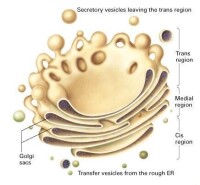
高爾基體是由數個扁平囊泡堆在一起形成的高度有極性的細胞器。常分佈於內質網與細胞膜之間,呈弓形或半球形,凸出的一面對著內質網稱為形成面(forming face)或順面(cis face)。凹進的一面對著質膜稱為成熟面(mature face)或反面(trans face)。順面和反面都有一些或大或小的運輸小泡,在具有極性的細胞中,高爾基體常大量分佈於分泌端的細胞質中。因其看上極像滑面內質網,因此有科學家認為它是由滑面內質網進化而來的。
細胞結構
高爾基體由兩種膜結構即扁平膜囊和大小不等的囊泡組成。其表面看上去極像光面內質網。扁平膜囊是高爾基體最富特徵性的結構組分。在一般的動、植物細胞中,3~7個扁平膜囊重疊在一起,略呈弓形。弓形囊泡的凸面稱為形成面,或未成熟面;凹面稱為分泌面,或成熟面。小液泡散在於扁平膜囊周圍,多集中在形成面附近。一般認為小液泡是由臨近高爾基體的內質網以芽生方式形成的,起著從內質網到高爾基體運輸物質的作用。糙面內質網腔中的蛋白質,經芽生的小泡輸送到高爾基體,再從形成面到成熟面的過程中逐步加工。較大的液泡是由扁平膜囊末端或分泌面局部膨脹,然後斷離所形成。由於這種液泡內含扁平膜囊的分泌物,所以也稱分泌泡。分泌泡逐漸移向細胞表面,與細胞的質膜融合,而後破裂,內含物隨之排出。不同細胞中高爾基體的數目和發達程度,既決定於細胞類型、分化程度,也取決於細胞的生理狀態。

對比
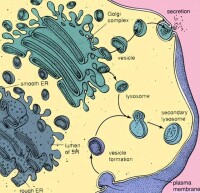
高爾基體是完成細胞分泌物(如蛋白)最後加工和包裝的場所。從內質網送來的小泡與高爾基體膜融合,將內含物送入高爾基體腔中,在那裡新合成的蛋白質肽鏈繼續完成修飾和包裝。高爾基體還合成一些分泌到胞外的多糖和修飾細胞膜的材料。
一、蛋白質糖基化
N-連接的糖鏈合成起始於內質網,完成於高爾基體。在內質網形成的糖蛋白具有相似的糖鏈,由Cis面進入高爾基體后,在各膜囊之間的轉運過程中,發生了一系列有序的加工和修飾,原來糖鏈中的大部分甘露糖被切除,但又被多種糖基轉移酶依次加上了不同類型的糖分子,形成了結構各異的寡糖鏈。糖蛋白的空間結構決定了它可以和哪一種糖基轉移酶結合,發生特定的糖基化修飾。
內部構造
二、參與細胞分泌活動
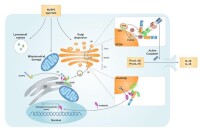
負責對細胞合成的蛋白質進行加工,分類,並運出,其過程是RER上合成蛋白質→進入ER腔→以出芽形成囊泡→進入CGN→在培養基(medial )Gdgi中加工→在TGN形成囊泡→囊泡與質膜融合、排出。
高爾基體對蛋白質的分類,依據的是蛋白質上的信號肽或信號斑。
細胞作用
三、進行膜的轉化功能
高爾基體的膜無論是厚度還是在化學組成上都處於內質網和質膜之間,因此高爾基體在進行著膜轉化的功能,在內質網上合成的新膜轉移至高爾基體后,經過修飾和加工,形成與運輸泡質膜融合,使新形成的膜整合到質膜上。
四、將蛋白水解為活性物質
如將蛋白質N端或C端切除,成為有活性的物質(胰島素C端)或將含有多個相同氨基序列的前體水解為有活性的多肽,如神經肽。
五、參與形成溶酶體
現在一般都認為初級溶酶體的形成過程與分泌顆粒的形成類似,也起自高爾基體囊泡。初級溶酶體與分泌顆粒(主要指一些酶原顆粒),從本質上看具有同一性,因為溶酶體含多種酶(主要是各種水解酶),是蛋白質與酶原顆粒一樣,也參與分解代謝物的作用。不同處在於:酶原顆粒是排出細胞外發揮作用,而溶酶體內的酶類主要在細胞內起作用。
六、參與植物細胞壁形成
高爾基體
高爾基體還有其他功能。如在某些原生動物中,高爾基體與調節細胞的液體平衡有關係。
1、順面膜囊 (cis Golgi) 接受來自內質網新合成的物質並將其分類后大部分轉入高爾基體中間膜囊,小部分蛋白質與脂質再返回內質網(駐留在內質網上)。其他如:蛋白的O-連接,跨膜蛋白的醯基化,日冕病毒的裝配也是在此。
2、中間膜囊(medial Golgi)多數糖基化修飾、糖脂的形成、多糖的形成。有很大的膜表面,增大了合成與修飾的有效面積。
3、反面膜囊(trans Golgi)ph比其他部位低。功能是蛋白質的分類與包裝以及輸出,“晚期”蛋白質修飾。並保證蛋白與脂質的單向轉運。
高爾基體反面膜囊網路結構(TGN)最近被發現可以參與炎症小體(又稱“炎性小體”,這裡的炎症小體的受體蛋白為NLRP3)響應外界信號進而組裝的信號軸。炎症小體是一個蛋白複合物,主要包含受體蛋白、接頭蛋白ASC以及下游的胱天蛋白酶caspase-1。炎症小體的種類和功能較為複雜,在這裡不作贅述。而對於NLRP3受體蛋白,其結構組成如下:
而下游的接頭蛋白ASC結構組成如下:
在受體蛋白被激活后可以通過自身的PYD結構域與接頭蛋白的PYD結構域結合以激活ASC,隨即後者通過自身的CARD結構域激活下游caspase-1,促使其自切割成2部分,進一步切割並促進細胞白介素等相關細胞因子的成熟。
在PYD和NACHT結構域之間,存在著高度保守的正電區域,其中僅含4個氨基酸的KKKK模體在所有已知的NLRP3基因的蛋白質中都至少含有3個正電氨基酸。當收到炎症小體激動劑的時候,TGN被發現可以解體形成囊泡狀結構(dTGN),這些結構上的負電磷脂PtdIns4P可以和NLRP3的正電荷結合,並促使後者形成聚合物,進而使NLRP3得以激活。
該機制在一定程度上 打破了TGN作為高爾基體固定組成部分的印象,開啟了高爾基體作為炎症反應信號軸“信使”的 新身份。
細胞中蛋白質的合成從細胞核中的基因組DNA轉錄合成信使RNA(mRNA)開始,mRNA穿過了細胞核到達核外,在內質網(ER)上合成了蛋白質,此時蛋白質會從內質網上以小囊泡的形式脫離下來,其目的地就是物流中心——高爾基體,就像工廠裡面生產出來的商品被輸送到物流中心再向用戶配送一樣。其實,被輸送到高爾基體的蛋白質上面已經標記了蛋白質的去向,就像商品進入物流中心時都會貼好收貨人地址一樣。此時,高爾基體就會讀取蛋白質的“收貨地址”,如果這個蛋白質上有“分泌”信號,高爾基體就知道這個蛋白質需要被分泌到細胞外,於是就形成一個可以分泌蛋白質到細胞外的小囊泡,把需要分泌到細胞外的蛋白質包裹到裡面並最終分泌到細胞外。如果蛋白上有信號表示該蛋白質應該留在內質網,高爾基體就會形成另外一種小囊泡,把蛋白送回到ER。高爾基體就是這樣通過閱讀蛋白質上的信息把蛋白運送到“收貨人”手中。高爾基體物流中心的功能還不止是這些,它還會對運輸的貨物——蛋白質進行加工,這個加工過程就是把各種寡糖鏈連接到蛋白上,這個過程稱為糖基化,而這種糖基化是蛋白最終可以執行各種功能的保證。
1、高爾基體肥大。高爾基體肥大見於細胞的分泌物和酶的產生旺盛時。巨噬細胞在吞噬活動旺盛時,可見形成許多吞噬體、高爾基複合物增多並從其上斷下許多高爾基小泡。
2、高爾基體萎縮。在各種細胞萎縮時可見高爾基體變小和部分消失。
3、高爾基體損傷。大多出現扁平囊的擴張以及扁平囊、大泡和小泡崩解。
基本信息
- 中文名
- 高爾基體
- 外文名
- Golgi apparatus, Golgi bodies
- 別名
- 高爾基複合體、高爾基器
- 拼音
- gaoerjiti
- 類型
- 細胞體
- 提出者
- 特點
- 高爾基體的形成面與順面具嗜鋨性
- 植物體內作用
- 與細胞壁形成有關
- 縮寫
- GA
- 動物體內作用
- 蛋白質的分選、加工,形成溶酶體