淡水生態
淡水生態
淡水生態指一定水域內所有生物(即生物群落)與它們的理化環境相互作用,通過物質循環和能流而構成的具有一定結構和功能的統一體。
目錄
━━━━━━━━━━━━━━━━━━━━━━━━
淡水生態系統
淡水環境
湖泊
水庫
池塘
河流
泉
淡水生物群落
淡水生物的生態類別
靜水生物群落
沿岸帶群落
敞水帶群落
深水帶群落
流水生物群落
急流帶群落
滯水帶群落
河道帶群落
養分循環和能流
養分循環
能流
演替與平衡
━━━━━━━━━━━━━━━━━━━━━━━━
研究內陸水體中生物與生物之間以及生物與環境之間的相互關係。包括生物個體、種群、生物群落和生態系統等層次,而生態系統已成為目前的研究重心。內陸水體包括江河、湖泊、水庫、池塘等,具有廣泛的地理分佈和懸殊的大小規模,其總面積約佔地球面積的0.5%。這些水體不僅是人類生活與工農業生產用水的主要來源,而且在航運、發電、漁業等方面給人類帶來了許多利益。同時,內陸水體作為生物圈的組成部分,對調節全球氣候、維持自然環境的穩定性起著重要的作用。淡水生態系統的研究,有助於闡明系統中的物質循環、能流以及演替和平衡的規律,為加強內陸水體的水質管理、防治水體污染或富營養化、合理開發和利用水體生物生產力提供科學依據。
淡水生態系統
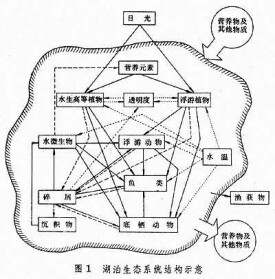
指一定水域內所有生物(即生物群落)與它們的理化環境相互作用,通過物質循環和能流而構成的具有一定結構和功能的統一體 (圖1)。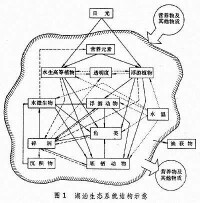 生態系統的非生物組分包括:氣候條件(溫度、光照及其他物理因素),參加物質循環的無機物質(碳、氮、磷、二氧化碳、水等),以及聯繫生物和非生物的有機化合物(蛋白質、碳水化合物、脂類、腐殖質等)。生態系統的生物組分依其生態功能可分為:生產者,主要指浮游植物和水生高等植物;大型消費者,包括浮遊動物、底棲動物、游泳動物等;以及微型消費者,主要是細菌和真菌。淡水生態系統的結構,不僅視靜水與流水兩大類型而異,而且同一類型的各種棲息地之間往往也有明顯的差別。
生態系統的非生物組分包括:氣候條件(溫度、光照及其他物理因素),參加物質循環的無機物質(碳、氮、磷、二氧化碳、水等),以及聯繫生物和非生物的有機化合物(蛋白質、碳水化合物、脂類、腐殖質等)。生態系統的生物組分依其生態功能可分為:生產者,主要指浮游植物和水生高等植物;大型消費者,包括浮遊動物、底棲動物、游泳動物等;以及微型消費者,主要是細菌和真菌。淡水生態系統的結構,不僅視靜水與流水兩大類型而異,而且同一類型的各種棲息地之間往往也有明顯的差別。
淡水生態
生態系統的生物組分通常分佈在空間的不同層次,它們之間主要以食物鏈(網)的形式相互聯繫。在生態系統的代謝過程中,綠色植物以日光為能源將無機物質合成可供消費者利用的有機物質,其食物潛能沿著不同的食物鏈逐級傳遞;死亡有機物質則被微型消費者分解,結果釋放出可被綠色植物重新利用的無機營養物。於是,生態系統中不斷地進行著營養物質在環境與生物之間的往複循環,以及能量從日光到生物的單向流動。物質循環和能流是生態系統的最重要的功能,而生物生產力是生態系統的這兩大功能的綜合表徵。
任何生態系統都是由簡單到複雜,從低級到高級不斷發展的。淡水水體發育和演替的基本模式,是從貧營養到富營養和由水體到陸地。生態系統的發育,一方面起因於系統內生物組分的活動,另一方面受外界因素的影響。在經過一定發育階段的生態系統中,各生物組分之間、群落與環境之間、以及結構與功能之間的相互關係是相對穩定和協調的,可通過信息反饋來維持自身的動態平衡。但生態系統的自我調節能力有限,其平衡狀態往往因強度過大的外來干擾而遭到破壞。20世紀60年代以來,淡水水體的嚴重污染或富營養化,都是人為因素所造成的自然生態平衡失調的實例。因此,加強淡水生態系統的管理,使之朝著有利於人類的方向發展,已成為關係到國計民生的一個重大問題。
淡水環境
地球上的水量共約137×108立方公里,其中96.5%的水積蓄在海洋,餘下的分佈在兩級冰冠、冰川、地下、內陸水體、土壤和大氣中。內陸水體的水量僅佔地球總水量的0.016%,其中淡水水體的水量佔55%,鹽湖和內陸海的水量佔45%。地球各部分的水量分佈,是通過降水、徑流和蒸發所構成的水循環而維持相對穩定的,內陸水體在這一循環過程中起重要作用。
內陸水體大體上可分為靜水水體(如湖泊、水庫、池塘)和流水水體 (包括泉、溪流及江河)兩大類。這些水體不僅地理位置和氣候條件不同,而且具有多種多樣的形態、結構和化學組成,因此生物的環境條件極為複雜。在淡水生態學研究中,水環境的溫度、光照(決定於透明度)、水流(尤其在河流中)、溶解氣體(氧和二氧化碳)、營養鹽類(主要是磷酸鹽和硝酸鹽)等,通常被認為是最重要的限制因素。
湖泊 靜水水體的主要類型。地球上可利用的淡水大部分貯存在湖泊中。從地質年代來看,湖泊的存在是短暫的,現有湖泊大都形成於較晚的歷史時期。如北歐、加拿大和美國北部地區的很多湖泊,大約在1~1.2萬年前最後的冰川退卻時形成。按照湖盆的成因,湖泊可分為構造湖、冰川湖、火山湖、山崩湖、溶解湖、河成湖、風成湖、海灣湖等類型。不同類型的湖泊具有不同的底質和形態特徵,而這些特徵直接關係到湖泊的理化環境。
湖泊作為生物的棲息地,通常可按光照條件分為沿岸帶、敞水帶和深水帶 3個不同的生境。沿岸帶指靠近湖岸的淺水區,日光可以直射到底,一般被水底長出的水生高等植物所佔據。沿岸帶以外從水面直到光的有效透射深度(即補償深度)為止的水層稱敞水帶。在所謂補償深度處光合作用放出的氧氣正好滿足呼吸作用的消耗,其光照強度通常為飽和光強度的1%。上述兩個區域都屬於光亮帶。敞水帶以下,即由補償深度直至水底的區域是深水帶,這裡光照微弱或無光線,不見大型植物分佈,但底部可有大量微生物滋生。淺水湖泊不存在深水帶,其他湖泊的敞水帶和深水帶都大於沿岸帶。一般說,湖泊的沿岸帶光照、氧氣和營養條件最佳,生物的種類和數量都很豐富。因此在水體生產力研究中常用湖岸線發育指數(DL)作為湖泊生產性能的一個指標。
湖泊中溫度(熱)、溶解氣體和營養鹽類的空間分佈主要取決於湖水的運動,由於引起湖水運動的主要因素(風和水的密度差)直接受氣候影響,因此湖中溫度、光照、養分等要素空間分佈的季節變化隨著湖泊的地理位置的不同而有很大差別。在溫帶地區,夏季的風力只能引起湖泊上層的水循環(湍流混合),致使湖中(除淺水湖泊外)出現溫度隨深度下降的正向分層現象。按照湖中的溫度梯度可將湖水分為湖上層、溫躍層和湖下層,其中溫躍層的溫度隨深度急劇下降(溫度通常每米可下降1℃)。冬季湖面結冰,冰層阻止湖中熱量的進一步散失,結果出現湖下層溫度高於湖上層的反向分層現象。春季和秋季分別是湖泊吸熱和放熱的時期,在強大的湍流混合和水密度差所引起的對流混合作用下,湖泊的上下水層出現循環(翻轉)而處於同溫狀態。熱帶湖泊溫度的季節變化很小,僅在某些年份於較冷的季節出現水循環,一般情況下保持著溫度梯度很小的正分層。根據湖泊的熱學特徵,可將世界上的大多數湖泊分為6個類型:二次混合湖(溫帶,水溫4℃左右);冷一次混合湖(兩極,水溫不超過4℃, 夏季循環);熱一次混合湖(溫帶或亞熱帶,水溫不低於4℃,冬季循環);寡混合湖(熱帶、很少混合);多次混合湖(赤道、高山地區,連續混合);和部分混合湖(由鹽類引起的永久性化學分層,水循環僅限於湖上層)。伴隨湖泊的熱分層,也出現氧分層現象。在溫帶湖泊夏季熱分層期間,湖中的溫躍層往往位於補償深度之下,結果湖下層因氧供應完全斷絕而陷於缺氧狀態,這種情況在富營養湖泊中更為嚴重。
湖泊的化學組成和營養狀況,因湖盆及其流域的地理和地貌特徵以及湖泊的發育過程而異。目前廣泛採用的世界湖泊分類方法有兩種:一是以生產力為依據的正常湖泊的貧營養-富營養系列;二是具有異常化學性質的特殊湖泊類型。湖泊的生產力或“肥力”取決於外源營養物、地質年齡和深度,大多數湖泊都可按初級生產力分類。貧營養湖泊一般較深,養分含量和初級生產力均低,處於地質學上的年輕階段。相反,富營養湖泊較淺,營養物含量和初級生產力均高,並且經歷了較長的地質年代。因此,在比較水色、透明度、底質、溶解氧、pH、無機和有機物含量,以及生物的組成和多度時,典型的貧營養湖泊與富營養湖泊往往是完全不同的。在貧營養-富營養系列中,按照給定指標的梯度,可以劃分出許多中間型湖泊,如貧-中營養湖、中營養湖、中-富營養湖等。特殊類型的湖泊主要包括:腐殖營養湖 (含高濃度腐殖酸),鹽湖(含鹽量高,分佈於乾旱氣候地區)和酸性或鹼性湖(分佈於活動的火山地區,受岩漿的影響大)。在這些湖泊中,化學條件嚴酷,通常僅有很少的具有特殊適應能力的生物。
水庫 近200年來,世界各地所建水庫的貯水量在內陸水體中佔有相當的比重。水庫的環境條件與天然湖泊有許多相似之處,上述有關天然湖泊的生境劃分和營養分類基本上適用於水庫。水庫的特點是水位不穩定和混濁度大,所以生物生產力往往低於天然湖泊。水庫的熱量、氧氣和營養物收支依其排水方式而異。如果水閘設在水壩底部,那麼排出的就是溫度低、營養物豐富和氧氣貧乏的水,而溫度高、營養物缺乏和氧氣充足的水留在水庫中。這不僅會使水庫失去必需的營養物,從而導致生產力降低,而且將對下游地區產生多方面的影響。若從水庫表面排水,其結果則與天然湖泊相同。
池塘 小型靜水水體,沿岸帶較大,而敞水帶和深水帶都較小或不存在, 故水體中的理化條件較為一致。一部分池塘是自然形成的(天然池塘,如河流改道后留下的部分舊河床、石灰岩底質被溶解所形成的窪地),或產生於某些動物(如河狸)的活動,但數量最多的是為了工農業用水而修築的人工池塘。池塘的水量變動較大,甚至有些池塘在一年中的部分時間內是乾涸的,這類池塘常稱臨時池塘。多數池塘都很肥沃,尤其是作為漁業水體的池塘,往往經人工施肥,因此常有較高的生產力。
河流 流水水體的主要類型,其貯水量約佔內陸水體總水量的 0.5%, 亦是地球上水循環的一個重要環節。與靜水水體不同,河流的首要特徵是具有連續的水流,這在很大程度上決定著河流中許多理化成分和小型生物的分佈。河水的流速決定於河床的坡降、深度、寬度和粗糙度,並且在很大程度上受排水量的影響。同一河流的不同部分和不同時期的流速常有很大的變化;在特定條件下,河流的某些區域可能處於靜水狀態。因此,河流中通常存在急流帶和滯水帶兩個不同的生境:前者指淺水區,流速大,底質堅硬;後者指深水區,水流緩慢,底質常鬆軟。某些河流的下游水流相當平緩,急流帶與滯水帶的差別基本消失,這部分河段常被稱為河道帶。河流的另一個特徵是深度和橫斷面顯著小於湖泊和水庫,具有較大的水-陸連接面,成為更開放的生態系統。實際上,大多數河流中有機物質主要來自陸地和鄰近的靜水水體,尤其是處於森林地帶和肥沃平原的河流接受外源物質更多。由於河流的深度小,與空氣的接觸面大,河水又不停地運動和混合,河流中的溶解氧通常較充足,並且幾乎沒有熱分層或化學分層現象。河流排水量的季節變動大,不同河段的流量差異顯著,外源物質的輸入具不確定性,所以河流的化學組成往往隨時間和距離而有很大的變化。同時,河流往往被看作有效的廢物處理系統,因而容易遭受有毒物質污染。
泉 地球上數量很少的一類小型流水水體,具有相對穩定的溫度、流速和化學組成。常見的幾種較為重要的泉是:溫泉(分佈於火山地區,通常含鹽量高)、硬水泉(分佈於石灰岩地區,具有所在地的平均溫度)和軟水泉(水從頁岩、沙岩和石英岩滲出)。泉流的理化性質固然在泉水流向下游的過程中發生變化,但是任一給定點的理化性質總是比較穩定的,因此提供了一個天然的生態梯度。鑒於某些大泉流中群落結構和代謝的穩定性,許多生態學家都把泉看作一個重要的研究領域。
淡水生物群落
生活在淡水棲息地的各類生物,通常分佈在不同的空間層次和佔據著各自的生態位,彼此之間存在著複雜的相互關係,這些生物種群的集合體被稱為生物群落。在生態學研究中,除了按水體或生境來研究整個生物群落外,還可以著眼於一些規模較小的群落單元,如浮游生物群落,底棲生物群落、周叢生物群落等。
淡水生物的生態類別 淡水生物依其在群落中所處的營養級可以分為:生產者(自養生物),主要包括浮游植物、水生高等植物和著生藻類,其次是一些光合細菌和化能自養細菌;大型消費者(牧食性和捕食性異養生物),指吃其他生物或有機碎屑的動物,主要有浮遊動物、底棲動物和各種魚類;微型消費者(腐食性異養生物或分解者),主要指細菌和真菌。淡水生物群落的每一個營養級上分佈著多種生物,它們在群落代謝中所起的作用很不相同,因此常有優勢種類與次要種類的區別。淡水生物依其生活方式或生活習性,可分為浮游生物、游泳動物、底棲生物、周叢生物和漂浮生物5個生態類群。
靜水生物群落 按照靜水棲息地的3個生境類型,靜水生物群落可分為沿岸帶群落、敞水帶群落和深水帶群落,這些群落的相對重要性主要決定於水體中3個生境的相對大小。一般說,沿岸帶是池塘的主要生產區,而湖泊和水庫的主要生產區則是敞水帶。
沿岸帶群落 沿岸帶的生產者主要包括底生植物和各種藻類,其次是漂浮植物。底生植物的分佈通常有明顯的水平成帶現象,即從岸邊到深處隨著水深的變化而出現3個不同的植物帶:①挺水植物帶,植物的莖葉高出水面,主要包括蘆葦、水燭、闊葉菰、蓮等;②浮葉植物帶,植物的葉浮在水面,主要有菱、荇菜、睡蓮等;③沉水植物帶,植物體完全沉沒在水下,主要種類為眼子菜科植物,其次有苦草、黑藻、等。藻類主要包括硅藻、綠藻和藍藻。除了一部分絲狀藻類和著生藻類依附於底生植物外,大部分藻類都為浮遊性。底生植物的分佈主要決定於水層的透明度,其生長情況常與浮遊藻類的生長呈負相關。在富營養化水體中,由於藍藻水華嚴重阻礙了光的透射,底生植物往往瀕於滅絕。漂浮植物主要有浮萍、滿江紅、水浮蓮等,在水面平靜的地方(如池塘和湖灣)常有大量分佈。
沿岸帶的消費者生物極為豐富,屬於5個生態類群的各級消費者都有存在。某些動物(尤其是周叢生物)的水平成帶分佈常與底生植物的分佈相平行,但有很多種類幾乎分佈在整個沿岸帶,並且動物的垂直成帶現象比水平成帶更為顯著。周叢生物主要有螺類、某些昆蟲幼蟲、原生動物、水螅、輪蟲、各種蠕蟲、苔蘚蟲等。底棲動物中以多種昆蟲幼蟲、環節動物(寡毛類)和軟體動物(螺、蚌)佔優勢。浮遊動物主要有大型的枝角類、橈足類的某些種類、介形類和輪蟲。很多甲殼動物都有笨重的外殼,常採用爬行方式運動,這是與敞水帶的種類明顯不同的特徵。漂浮動物通常僅有幾種水面昆蟲,如鼓甲、黽蝽等。游泳動物主要有多種昆蟲(幼蟲和成蟲)、兩棲類(蛙)、爬行類(龜、蛇)和各種魚類。其中魚類來往於沿岸帶與敞水帶之間,常在沿岸帶覓食和繁殖。
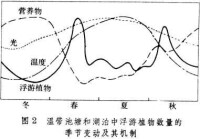
敞水帶群落 生產者包括浮游植物和一些浮遊自養菌。浮游植物主要有甲藻、硅藻、綠藻和藍藻,個體小,但生產力相當高。在北溫帶湖泊中,隨著溫度、光照和營養物再生的季節變化,浮游植物於春秋兩季各出現一次高峰(水華),夏冬兩季則處於較低水平(圖2)。春季水華主要由硅藻所引起,而秋季水華通常歸因於藍藻。
淡水生態
深水帶群落 深水帶不存在生產者,消費者種類也不多,其食物供應依賴於沿岸帶和敞水帶。群落的主要成員是細菌和真菌,它們所提供的再生營養物常通過水流和游泳動物帶到其他區域。大型消費者主要有搖蚊幼蟲、顫蚓、球蜆和幽蚊幼蟲。前3種屬於底棲動物,最後1種是部分浮遊動物,這些動物都有適應缺氧環境的能力。
流水生物群落 流水生物群落可按河流所在地的3類生境分為急流帶群落、滯水帶群落及河道帶群落。河流上下游的環境條件差別顯著,河流生物的分佈有明顯的縱向成帶現象,這點與靜水生物的水平成帶分佈不同。
急流帶群落 由典型的河流生物組成,主要是一些著生藻類和各種昆蟲幼蟲。生活在這裡的動物都具有特化的形態結構,明顯地適用於流水環境。平常見到的浮游生物來自鄰近的靜水水體。
滯水帶群落 生產者有絲狀藻類和一些沉水植物。消費者主要包括一些穴居或底埋動物和多種魚類,後者為了覓食亦常在急流帶出現。在某些流速很低的河流中還生長著浮游生物。
河道帶群落 其結構類似於靜水生物群落。除河流生物外還可見到很多靜水生物。但由於河床底質的不均勻性,底棲動物通常以成團的形式分佈。
養分循環和能流
養分循環 淡水生態系統是生物圈的一個組成部分,一方面與大氣保持著連續的氣體交換,另一方面通過徑流與陸地、農田和鄰近的其他水體存在著廣泛的聯繫。因此,淡水生態系統中的養分處於經常的變動狀態,其輸入與輸出關係依水體的類型、形態結構和地理分佈而異。大多數靜水水體排出的是養分貧乏的表面水,所以養分的輸出明顯地小於輸入,水體有向富營養化發展的趨勢。
雖然水體可由多種途徑接受一定的外源養分,但是經常的和大量的養分供應主要依賴於系統內部的循環。這一循環的基本過程是:綠色植物以日光為能源,由無機養分合成有機物質,供消費者利用,同時通過生物的呼吸和死亡有機物質的分解,養分重新釋放出來。
淡水生態系統中有機物質的分解,是生物的和非生物的因素共同作用的結果,但是一般起主要作用的是以這些有機物質為食物來源的腐食者生物。水體中的有機物質,一部分是系統內的動植物屍體和排泄物,另一部分是來自其他系統(外源)的有機物質,包括人類排放的污水和施入的有機肥料。在有機物質分解的過程中,各種異養微生物通常是同時或交替起作用的,任何一個分解者種群都不能單獨實現把有機物質完全分解的任務。現知,一些小型的腐食性或碎屑食性動物,如原生動物、線蟲、介形類等,在有機物質分解過程中所起的作用很大。在某些生態系統中,小型動物對有機物質的分解甚至比細菌或真菌更為重要。
微生物的分解過程只是營養物再生的途徑之一,而且在某些情況下還處於從屬地位。例如,在浮游生物佔優勢而以牧食食物鏈為主要能流途徑的生態系統中,直接從浮遊動物的排泄物和分泌物中返回的養分,常比經微生物分解所得到的量高得多。研究表明,浮遊動物的排泄物中含有許多可溶性的無機和有機化合物,這些養分能直接被浮游植物所利用。細菌在分解植物組織時會將所獲得的磷酸鹽和硝酸鹽保留在體內,只有當它們被食碎屑的動物消費后營養物才能釋放出來。
在淡水生態系統中,養分的循環過程不是均衡的,而是存在著養分的吸收與釋放的周期變化。經32磷示蹤試驗發現,湖泊中大部分的磷或存在於生物體內,或被結合在沉積物中,而水中的溶解磷至多只有10%。因此,沉積物中養分的釋放率,在很大的程度上反映了養分循環的速度。由於不同水體的形態結構和理化狀況不同,沉積物中養分的釋放率往往差別很大。一般情況下,小型淺水湖泊沉積物中磷的周轉時間都較短,並且湖中往往出現養分的“短路代謝”,所以通常生產力較高。根據水體中物質循環的規律,努力使系統中的養分得到充分的循環和利用,是提高漁業水體生產力的一項重要措施。中國正在推廣的桑基魚塘,用魚塘底泥作為桑田肥料,桑葉養蠶,蠶砂(糞)用於施肥養魚。這在調節營養物的循環過程,充分利用自然資源和維護自然生態平衡方面,已取得良好的效果。(見農業生態)
能流 按國際生物學規劃(IBP)的廣泛調查結果,世界湖泊的總初級生產力變動在500千卡/(米2·年) (北極湖)和1萬千卡/(米2·年) (某些熱帶湖泊)之間。河流生態系統的初級生產力較低,除某些泉流外,通常很少超過1000千卡/(米2·年)。據估計,世界湖泊與河流的總凈初級生產量為每年0.6×1016千卡,占生物圈總凈初級生產量的0.82%,平均凈初級生產力為每年每平方米2250千卡。
淡水生態系統中的能流通過牧食食物鏈、捕食食物鏈和碎屑食物鏈共同實現。但在不同的水體中,這兩類能流途徑所起的作用往往有明顯差別。大型湖泊和水庫中以牧食食物鏈為主,而碎屑食物鏈在水生高等植物繁茂的水體中起主導作用。在後一類水體中,90%的初級生產量通過碎屑路線被利用。水體中的碎屑,除了來自死亡的動植物體之外,植食性動物的排糞是另一個重要來源。
食物鏈原理對淡水漁業生產有重要的指導意義。漁業生產的基本目標,是充分利用水體的餌料生物資源,最大限度地提高魚產量。為此,應採取一系列措施來調節水體的生產過程,使系統中的能源物質儘可能轉化為魚產品。這些措施包括:①多放養食物鏈短的魚類,以有效地利用生產者所固定的能量;②控制兇猛魚類種群的發展,以減少能量多環節傳遞所造成的損失和對放養魚類的危害;③注意天然雜食性魚類的繁殖保護,並適當增加食碎屑魚類的放養量,以提高水體中碎屑資源的利用率。目前,以鰱、鱅為主體的多品种放養,已成為中國淡水漁業生產上成功的經營方式。
演替與平衡
生態系統的發育,即通常所講的生態演替,指一定區域內連續進行的前一群落被后一群落所替代的發展過程。演替起因於系統內生物導致的環境改變、外來種的入侵等生物學過程,或起因於氣候變遷、土壤侵蝕、河流泛濫、環境污染等外部力量,也可能是這兩方面因素共同作用的結果。演替過程中依次出現的一系列群落構成一個演替系列,其中早期出現並且存在時間比較短暫的群落屬於先鋒階段,相繼出現的群落屬於發育階段或演替階段,而最終形成的穩定系統稱為演替頂極。演替是群落髮育的有序過程,這一過程基本上有一定的規律,因此通常可以預測。
淡水生態系統發育的基本模式,是從貧營養到富營養和由水體到陸地。這一過程通常可用湖泊的發育史來說明。湖泊形成初期,養分含量很低,常常僅有一些浮游生物。由於外源物質的輸入,湖中有機物質含量增高,並出現沿岸沉積物。於是,水生高等植物慢慢發展起來,水生昆蟲、環節動物、魚類等多種動物陸續遷入。隨著時間的推移,水底沉積物不斷增加,沿岸帶植物逐漸向湖心發展。因此,湖泊逐漸由深變淺,由大變小,直至整個水體最後完全消失。淡水生態系統的發育過程一般比陸地生態系統緩慢,並且往往因外源物質的大量輸入而出現十分複雜的情況。同時,在發育過程中,生態系統內一些較小的群落單元,如浮游生物群落、微生物群落等,常常表現明顯的季節演替現象。
生態系統是天然的控制系統,其自我調節依賴於負反饋機制。在淡水生態系統中,許多反饋路線存在於生物與生物和生物與環境之間,通過反饋,種群大小得以維持在一定的限度內。例如,當某種兇猛魚或浮游植物種群增長得過大,以致其食物或養分供應不能滿足時,就會出現大量的自然死亡,種群繁殖率下降,以及兇猛魚殘害其幼體的現象,結果種群數量傾向於減少。除捕食和競爭外,生態系統中所發生的其他相互影響,如寄生、共生等,都有維持系統相對穩定的趨向。
然而,生態系統的自我調節能力是有限度的,即使是處於成熟階段(即演替頂極)的生態系統,也只能承受一定強度的外來干擾(壓力)。當外部壓力超過所允許的限度時,就會破壞生態系統的平衡狀態,甚至造成整個系統的崩潰。工業污染物傾瀉入湖造成的湖泊生物大量死亡和湖泊生態系統的破壞(見污染生態)以及外源養分的輸入過多所引起的湖泊富營養化,都是自然生態平衡遭到破壞的典型實例。
大量的外源養分(磷、氮等)進入湖泊后,加速了藻類(主要是藍藻)的生長,水體中的有機物質大幅度增加。可是,藍藻不能很好地被植食性動物所利用,以致水體中出現初級生產的相對過剩。同時,大量的有機物質沉積於水底,其分解造成湖下層嚴重缺氧。結果,很多水生動物逐漸消失,而藻類生物量越來越大,水質不斷惡化。在富營養化水體中,由於群落結構的破壞和一系列功能失調,物質循環和能流過程不能正常完成。
外源物質人為地大量輸入所引起的水體富營養化,與天然情況下的水體富營養化根本不同。後者是在系統內生物學過程起主導作用的情況下產生,其變化過程是漸進的,漫長的,反映了生態演替的正常趨向。而人為的水體富營養化,則是在外部力量的影響下產生的;由於系統內的生物學過程受到嚴重干擾,因而生態演替的正常趨向發生逆轉。在人為的富營養化水體中,物種的多樣性明顯下降,采 r型策略的物種重新佔優勢,這可看作典型的演替逆轉現象。
20世紀70年代以來,由於人類經濟活動的影響,大量的淡水水體出現污染或富營養化,致使人類的生活和生產用水受到嚴重威脅,淡水生物資源遭到極大的破壞。因此,加強對淡水生態系統的保護,採取有效措施防治水體污染或富營養化,已成為關係到人類的生存和社會經濟發展的一個重大問題。在水體富營養化的治理中,通過採取污水截流、換水、挖泥等措施,已在不同程度上收到了改善水質的效果。實踐證明,只要減少或停止外源養分的輸入,就可以大大延緩或終止水體富營養化的進程。
參考書目
R.K.Barnes, K.H. Mann,Fundamentals of Aquatic Ecosystems, Blackwell Scientific Publications, Oxford,1980.
E.P.Odum,Fundamentals of Ecology, 3rd ed.,W.B.Saunders Co.,Philadelphia,1976.
基本信息
- 性質
- 生物圈的一個組成部分
- 外文名
- Freshwater Ecology