致病疫霉
致病疫霉
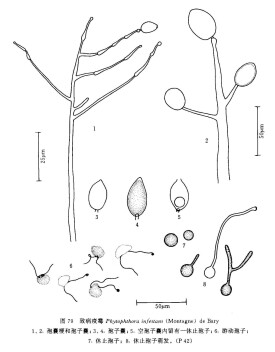
在馬鈴薯塊莖上,菌絲無色無隔,壁薄,自由分枝,粗4.9-12.0μm;未見菌絲膨大體。孢囊梗由菌絲生出,直立,無色,合軸分枝,粗5-9μm,頂端稍膨大並著生孢子囊,孢子囊成熟時被推向一側,梗繼續生長,於頂端再形成孢子囊,梗與孢子囊接觸點膨大,使整個孢囊梗呈粗細相間的節狀。孢子囊卵形、長卵形、橢圓形或檸檬形,具半乳突,頂生,24-54μm ×19-30μm,平均35.6μm×22.6μm [在固體培養基上為25-47μm×18-37μm(平均35.2μm× 23.5μm)],長寬比值為1.5,基部逐漸變細,脫落,柄短,柄長1.5-3.0μm;萌發產生遊動孢子或芽管,每一孢子囊可釋出5-12個遊動孢子。
休止孢子球形,直徑9.8-12.8(平均11.37)μm。厚垣孢子和有性器官均未見。
最低4-5℃,最適19-21℃,最高29℃。不能利用症粉。
中國分佈:番茄Lycopersicon esculentum Mill. :北京[PZ 43,P86(A1)]。
蜀羊泉Solanum lyratum Thunb.:四川雅安(HMAS 43729)。
據戴芳瀾(1979),此菌在中國台灣還為害十萼茄Solanum biflorum Lour,和茄S. melongena L.,後者在吉林也有報道。何漢興(Ho,1990)和蔡雲鵬(1991)報道在台灣為害龍珠Tubocapsicum anomalum(Franch et Sav.)Makino、番茄和馬鈴薯。
世界分佈:土耳其、馬來西亞、馬爾他、馬達加斯加、中國、烏拉圭、烏干達、日本、比利時、模里西斯、巴勒斯坦、巴西、巴拿馬、丹麥、牙買加、厄瓜多、以色列、印度、印度尼西亞、古巴、尼加拉瓜、加拿大、匈牙利、冰島、西班牙、百慕大群島、多米尼加、瓜地馬拉、阿爾及利亞、多巴哥、克里特島、芬蘭、希臘、阿根廷、蘇丹、留尼汪島、坦尚尼亞、英國、法國、波蘭、羅馬尼亞、波多黎各、委內瑞拉、埃及、南非、辛巴威、突尼西亞、保加利亞、挪威、南斯拉夫、宏都拉斯、美國、玻利維亞、莫三比克、夏威夷、荷蘭、特立尼達、哥斯大黎加、哥倫比亞、秘魯、菲律賓、薩爾瓦多、俄羅斯、喀麥隆、斯里蘭卡、葡萄牙、捷克、斯洛伐克、智利、奧地利、塞普勒斯、義大利、瑞典、瑞士、紐西蘭、蒙特塞拉特、摩洛哥、澳大利亞、墨西哥、德國。
致病疫霉Phytop有明顯的生理分化現象,變異性頗大。 “致病疫霉生理小種國際命名法規”用由4個基因型(R1R2R3R4)所決定的16個抗病品種作鑒別寄主,區分出16個生理小種。目前又陸續鑒定出11個R基因,相應出現更多的生理小種(Ribeiro,1978)。
致病疫霉最初被引入東半球可能是隨觀賞或藥用茄科植物上的卵孢子而傳帶的。此菌可能原產於墨西哥,因為它發生在當地的野生植物上,這些寄主植物種的大多數是抗病的或免疫的;很少發生在栽培的馬鈴薯上。P. in- festans起源於南美的秘魯或智利的說法也不可全信,因為那兒缺乏抗病品種。
基本信息
- 中文名
- 致病疫霉
- 拉丁學名
- Phytophthora infestans
- 界
- 真菌界
- 亞門
- 鞭毛菌亞門(Basidiomycotina)
- 門
- 真菌門(Eumycota)
- 綱
- 卵菌綱 Basidiomycetes
- 生長溫度
- 4-29℃,最適19-21℃
- 分佈
- 北京[PZ 43,P86(A1)]
- 形態特徵
- 休止孢子球形,直徑9.8-12.8μm