生殖細胞
生殖細胞
生殖細胞(germ cell),又稱配子(gamete),是多細胞生物體內能繁殖後代的細胞的總稱,包括從原始生殖細胞直到最終已分化的生殖細胞(精子和卵細胞),均為單倍體細胞,其中包含一條性染色體。此術語由A·恩格勒和K·普蘭特爾於1897年提出以與體細胞相區別。體細胞最終都會死亡,只有生殖細胞有延存至下代的機會。物種主要依靠生殖細胞而延續和繁衍。長期的自然選擇使每一種生物的結構都為其生殖細胞的存活提供最好的條件。
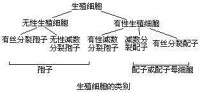
生殖細胞可以分成孢子和配子兩類:
是不需配合的生殖細胞,通常是無性的,可由減數分裂或有絲分裂產生,見之於原生動物中的孢子蟲綱和植物中。
是需經配合成合子後方能發育的生殖細胞,也稱性細胞,由減數分裂或有絲分裂產生。產生配子的細胞稱配子母細胞。這是未分化的原始生殖細胞,可在雄性或雌性生殖腺中分別分化為精子和卵。在低等植物中,配子母細胞在大多數情況下直接構成稱作配子囊的性器官。例如在原植體植物(Thallophyte)中的同形配子囊產生同形配子;異形配子囊產生異形配子等。
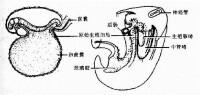
原始生殖細胞產生示意圖
極質,其中富含核糖核酸(RNA)的小顆粒叫極顆粒。經過受精、卵裂,含有極顆粒的細胞稱極細胞,就是果蠅的原始生殖細胞。如果將原在後部的極質注射到卵的前部,可使預定發育為體細胞的細胞發育為生殖細胞。
生殖細胞的類別
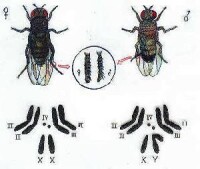
昆蟲的生殖細胞
由於有了對細胞的這種新的解釋,重新考慮受精過程的時機已經成熟。如果軀體的一切部位都由細胞組成,則性腺(卵巢和睾丸)是否也是如此?雄性和雌性的“種子物質”究竟是什麼?它是否也由細胞構成?雌、雄性的性細胞有什麼區別?這一類早已構思過的問題當然並不是一開始就提了出來,然而這些問題必然是細胞學說的邏輯結果。後來越來越明顯只有等到闡明了受精過程中細胞的作用,真正有生命力的遺傳學說才能建立。正是在這幾十年中生殖細胞這個概念才脫穎而出。
精子的獲能
精子中的半數含Y染色體(23,Y),半數含X染色體(23,X)。射出的精子雖有運動能力,卻無穿過卵子周圍放射冠和透明帶的能力。這是由於精子頭的外表有一層能阻止頂體酶釋放的糖蛋白。精子在子宮和輸卵管中運行過程中,該糖蛋白被女性生殖管道分泌物中的酶降解,從而獲得受精能力,此現象稱獲能(capacitation)。精子在女性生殖管道內的受精能力一般可維持1天。
卵子的成熟
從卵巢排出的卵子處於第二次成熟分裂的中期,並隨輸卵管傘的液流進入輸卵管,在受精時才完成第二次成熟分裂。若未受精,於排卵后12~24小時退化。
受精
動、植物受精作用的實質是父本和母本的生殖細胞(配子)互相融合,這兩個配子在形成新的合子上各自作出了同樣貢獻,而且關鍵過程是兩個配子的細胞核相互融合,這些觀點到了1844年左右已經逐步確立並被有關學者普遍接受。人們的注意力便開始轉移到細胞核上。細胞核是不是就像後生論者所設想的,僅僅是一團無定形的胚樣物質、也許只是在融合時才激發了卵細胞的發育過程?或者是細胞核雖小,卻具有嚴密結構,這肉眼不可見的顯微結構是否就是受精作用之後的一切非常精確並具有特異性的發育過程的關鍵?如果把細胞核僅僅看作是細胞發育和細胞分裂的引發物,就會認為它在完成了這一任務后就會被溶解掉,在新的細胞分裂之前或至少是在配子形成之前再重新形成。由於19世紀後半期的細胞學家所接受的都是生理學家或胚胎學家的教育訓練,他們的側重點是發育問題,因而用不著關心細胞核的連續性。他們很少過向性狀是怎樣從親代傳遞到子代的遺傳學問題。
植入
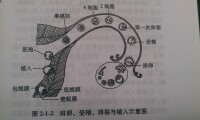
生殖細胞
胚泡的植入部位通常在子宮體和底部,後壁多於前壁。在內分泌失調,輸卵管炎症,粘連,狹窄等因素影響下,使胚泡未能進入子宮而在子宮以外的部位植入,發生宮外孕(ectopic pregnancy),常發生在輸卵管,偶也見於卵巢表面、子宮闊韌帶、腸系膜等處。