共找到2條詞條名為β-內醯胺酶的結果 展開
- β-內醯胺酶
- 青霉素酶
β-內醯胺酶
β-內醯胺酶
金屬β-內醯胺酶,屬Bush分類3群,Ambler分類B類,該群酶最大特點是可以水解青霉素類等抗生素,而對哌拉西林和氨曲南影響較小,其活性不被克拉維酸等β-內醯胺酶抑製劑所提高,但可被乙二胺四乙酸(EDTA)所抑制。酶活性中心需金屬鋅離子的參與,故稱為金屬β-內醯胺酶。
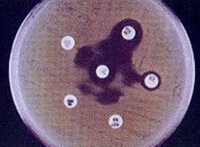
β-內醯胺酶
第一個報道的金屬酶是從蠟樣芽孢桿菌(Bacill us cereus)中發現的,該酶為鋅依賴酶。20世紀80年代初期日本從嗜麥芽窄食單胞菌中鑒定出第二種鋅依賴青霉素酶L1型酶,隨後又從嗜水氣單胞菌和脆弱擬桿菌中鑒定出多種能水解亞胺培南的金屬酶。這些酶都由染色體基因編碼。該類金屬酶分佈在蠟樣芽孢桿菌、嗜麥芽窄食單胞菌、脆弱擬桿菌、氣單胞菌屬和戈氏軍團菌中,除嗜麥芽窄食單胞菌外,在臨床上都極為罕見,而且都是單株散發的。
1991年日本學者在銅綠假單胞菌中發現了第一種質粒介導的金屬酶(IMP21),不久又從脆弱擬桿菌中發現了一種可轉移金屬酶,這兩個酶的發現意味著金屬酶已經從單株散發向隨機分佈過渡。現在已報道了10多種可轉移金屬酶: IMP21~8和VIM21~3,分佈在銅綠假單胞菌、不動桿菌和腸桿菌科細菌中,地域分佈上已經不再局限於日本,現已分佈至亞洲、歐洲和美洲的多個國家。
各種β-內醯胺類抗生素的作用機制均相似,都能抑制胞壁粘肽合成酶,即青霉素結合蛋白從而阻礙細胞壁粘肽合成,使細菌胞壁缺損,菌體膨脹裂解(胞壁粘肽合成過程見三十七章)。除此之外,對細菌的致死效應還應包括觸發細菌的自溶酶活性,缺乏自溶酶的突變株則表現出耐藥性。哺乳動物無細胞壁,不受β-內醯胺類藥物的影響,因而本類葯具有對細菌的選擇性殺菌作用,對宿主毒性小。近十多年來已證實細菌胞漿膜上特殊蛋白PBPs是β-內醯胺類葯的作用靶位,PBPs的功能及與抗生素結合情況歸納於。各種細菌細胞膜上的PBPs數目、分子量、對β-內醯胺類抗生素的敏感性不同,但分類學上相近的細菌,其PBPs類型及生理功能則相似。例如大腸桿菌有7種PBPs,PBP1A,PBP1B與細菌延長有關,青霉素、氨苄西林、頭孢噻吩等與PBP1A、PBP1B有高度親和力,可使細菌生長繁殖和延伸受抑制,並溶解死亡,PBP2與細管形狀有關,美西林、棒酸與硫黴素(亞胺培南)能選擇性地與其結合,使細菌形成大圓形細胞,對滲透壓穩定,可繼續生幾代后才溶解死亡。PBP3功能與PBP1A相同,但量少,與中隔形成,細菌分裂有關,多數青霉素類或頭孢菌素類抗生素主要與PBP1和(或)PBP3結合,形成絲狀體和球形體,使細菌發生變形萎縮,逐漸溶解死亡。PBP1,2,3是細菌存活、生長繁殖所必需,PBP4,5,6;與羧肽酶活性有關,對細菌生存繁殖無重要性,抗生素與之結合后,對細菌無影響。
β-內醯胺酶可能添加的食品:乳製品,特別是“無抗奶”。β-內醯胺酶類物質被用作牛奶中抗生素的分解劑。
解讀β-內醯胺酶:不得不先說說β-內醯胺類抗生素。β-內醯胺類抗生素是在牛乳生產中應用最廣泛的抗生素,用於治療牛乳腺炎和其他細菌感染性疾病。按照國家規定,使用抗生素藥物后一定時間內的乳汁,不得作為供人食用的原料。同時國家在《生鮮牛乳收購標準》中規定,生鮮乳中不得檢出抗生素。然而就中國奶牛飼養環境而言,牛奶的絕對“無抗”較難達到,針對這種情況,市場上出現了“抗生素分解劑”,該分解劑可選擇性分解牛奶中殘留的β-內醯胺抗生素,其成分就是β-內醯胺酶。
β-內醯胺酶(金玉蘭酶製劑),可能添加到乳與乳製品中,起到掩蔽抗生素的作用,但是由於該製劑的安全性風險未知,因此所有乳製品生產企業嚴禁在產品中添加此類物質。
基本信息
- 中文名
- β-內醯胺酶
- 外文名
- Beta Lactamase
- CAS登錄號
- 9073-60-3