內共生學說
內共生學說
內共生學說(endosymbiotic theory)關於線粒體起源的一種學說。認為線粒體來源於細菌,即細菌被原核生物吞噬后,在長期的共生過程中,通過演變,形成了線粒體。該學說認為:線粒體祖先原線粒體(一種可進行三羧酸循環和電子傳遞的革蘭氏陰性菌)被原始真核生物吞噬后與宿主間形成共生關係。在共生關係中,對共生體和宿主都有好處:原線粒體可從宿主處獲得更多的營養,而宿主可借用原線粒體具有的氧化分解功能獲得更多的能量。但內共生學說無法解釋細胞核起源。
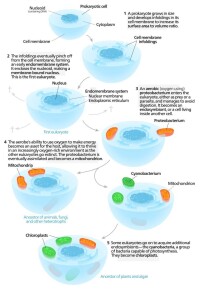
共生起源假說
1905年,Konstantin Mereschkowsky最先提出葉綠體是由原先的內共生體形成的這一想法,隨後1920年代Ivan Wallin提出了對線粒體的相同想法。隨著人們發現它們含有DNA,這些想法被Henry Ris重新提出。
內共生假說被琳·馬古利斯所普及。在她1981年的《細胞進化中的共生》中,她認為真核細胞起源於相互作用的個體組成的群落,也包括內共生的螺旋體形成了真核生物的鞭毛和纖毛。后一種想法沒有被廣泛接受,因為鞭毛缺乏DNA,在結構之外和原核生物也沒有明顯相似性。根據Margulis和Sagan(1996),“生命並不是通過戰鬥,而是通過協作佔據整個全球的”,而達爾文關於進化由競爭驅動的想法是不完善的。然而也有人認為內共生更像奴隸制而非互利共生。
有人認為過氧化物酶體可能也具有內共生起源,儘管它們缺乏DNA。 Christian de Duve認為他們也許是最早的內共生體,使得細胞能夠抵抗地球大氣中越來越多的氧氣。然而,現在看來它們可能是“從頭合成”的,而非具有共生起源。
這一假說由於證據充分,已被越來越多的人所接受。主要證據如下:
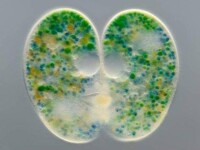
纖毛蟲(Nassula ornata)與藻類的共生現象
(2)葉綠體和線粒體都有其獨特的DNA,可以自行複製,不完全受核D NA的控制。線粒體和葉綠體的DNA同細胞核的DNA有很大差別,但同細菌和藍藻的DNA卻很相似。藍藻的核糖體RNA(rRNA)不僅可以與藍藻本身的DNA雜交,而且還可與眼蟲葉綠體的DNA雜交,這些都說明它們之間的同源性。
(3)線粒體和葉綠體都有自己特殊的蛋白質合成系統,不受核的合成系統的控制。原核生物的核糖體由30S和50S兩個亞基組成,真核生物的核糖體由40S和60S兩個亞基組成。線粒體和葉綠體的核糖體分別與細菌和藍藻的一致,也是由30S和50S兩個亞基組成,這說明細菌和線粒體、藍藻和葉綠體是同源的。抗生素可以抑制細菌和藍藻的生長,也可以抑制真核生物中的線粒體和葉綠體的作用,這也說明線粒體與細菌、葉綠體與藍藻是同源的。
(4)線粒體、葉綠體的內、外膜有顯著差異,內、外膜之間充滿了液體。研究發現,它們內、外膜的化學成分是不同的。外膜與宿主的膜比較一致,特別是和內質網膜很相似;內膜則分別同細菌和藍藻的膜相似。
1.內共生過程中,擁有先進的氧化代謝途徑的好氧細菌無疑應該佔優勢,但按內共生假說,好氧細菌反而逐步喪失了獨立自主性並將其遺傳信息成批地轉移到了宿主細胞中,這種觀點不符合進化論思想。
2.好氧細菌據估計發生在15億年以前(甚至更晚一些),但化石提供的證據說明藍藻大約在22-27億年以前就形成了。那麼,為什麼藍藻開始共生的時間卻遠比好氧細菌進行共生的時間遲得多呢?
4.在葉綠體基因與線粒體基因中發現內含子是驚人的,因為細菌基因中沒有發現內含子。這種內含子的來源及其對生物的效應目前沒有得到很好的解釋。
5.線粒體含有肌動蛋白-肌球蛋白系統,但細菌和藍藻中沒有這種系統,也沒有相應的運動能力。
6.內共生假說沒有很好解釋細胞核的進化。真核生物的C值(基因組大小)比原核生物高約3.5個數量級,如果兩個原核細胞通過吞噬進行共生,為何會出現如此巨大的基因組差異?
7.藍藻可以進行有氧呼吸,為什麼植物還需要線粒體?

基本信息
- 中文名
- 內共生學說
- 外文名
- endosymbiotic theory
- 共生
- 是生物界的普遍現象
- 1970年
- 《真核細胞的起源》書中正式提出
- 1905年
- 提出葉綠體是原先的內共生體形成
- 認為
- 線粒體來源於細菌