嵌合體
嵌合體
遺傳學上用以指不同遺傳性狀嵌合或混雜表現的個體,亦指染色體異常類型之一。有時也有同一器官出現不同性狀的生物體的意思。出現該病狀的人員往往伴隨精力極度旺盛,有時出現多重人格的精神錯亂癥狀。目前臨床還出現偶爾患者血型變換的癥狀。

2、染色體異常類型之一。來自不同合子的細胞系所組成的個體。又稱異源性嵌合體。起源於同一合子發育成不同核型的細胞系所形成的個體則稱同源性嵌合體(mosaic),又稱鑲嵌體。在動物中常見的嵌合體有牛弗里馬丁(freemartin) ——生殖器不全牝犢。即一雄一雌雙胎而產生的小母牛,其核型可出現性染色體嵌合,如60,XX/XY。
3、同一器官出現不同性狀的生物體,這是由於體細胞突變形成遺傳型不同的細胞所組成的。植物組織移植也會產生嵌合體。
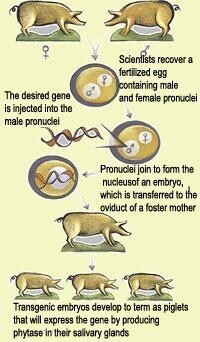
4、動物定向基因轉移技術中,通過向囊胚腔注射被外源基因轉化了的胚胎幹細胞,使得發育成為的個體中含有不同基因型的細胞,產生的個體也叫嵌合體,即該生物體中嵌合了兩種不同遺傳結構的細胞(一種是基因型被改變了的細胞,另一種是原來的基因型的細胞)。動物轉基因技術中利用嵌合體(可視為雜合體)之間的交配,產生純合體,該純合體即為轉基因動物,其兩條染色體上的某個基因位點都是經過人工改造的。
1646年義大利園藝學家費拉里烏斯首先報道了自然發生的柑橘類嵌合體;1860年德國養蜂學家奧伊格斯特發現蜜蜂中存在嵌合體;1875年C.R.達爾文把不同種的接穗和砧木產生的中間狀態的植物稱為營養雜種;1910年德國遺傳學家E.鮑爾證明:所謂營養雜種其實是嫁接嵌合體;1907年德國植物學家和遺傳學家H.溫克勒把嵌合體比喻為希臘神話中的獅首羊身蛇尾的神獸,這就是chimera一詞的由來;1929年美國遺傳學家A.H.斯特蒂文特利用果蠅的嵌合體研究胚胎髮育;1977年P.S.卡爾森等報道利用組織培養的方法培養出煙草種間嵌合體,為植物遺傳工程提供了新的可能性。2007年美國發現“半同卵性雙胞胎”,這是個同時擁有卵巢和睾丸組織的小孩,即是雌雄同體的“陰陽人”
嵌合體在果樹園藝中有很高的經濟價值。例如日本土橋紅蜜柑是扇形嵌合體,在紅色果皮上鑲嵌有艷麗的黃色條紋。許多名貴的花卉也是嵌合體。嵌合體一般不能通過有性生殖保種,但可以採用組織培養方法保種和繁殖。果蠅的雌雄嵌合體及異表型小鼠是行為遺傳學和發生遺傳學研究的材料。
嵌合體的詞意是古希臘神話中的吐火獸(chim-aere, chimera),具有羊身蛇尾的怪物。在生物學中的嵌合體是指同一個體中不同基因型的組織相互接觸而存在的現象,動物中通常把同樣的情況稱為鑲嵌(mosaic)。
嵌合成分來自原屬同一受精卵的嵌合體,大部分由染色體畸變和基因突變(自發或誘發)產生。可分為兩種:
嵌合體
基因嵌合體
①胞質基因嵌合體,如常春藤、紫茉莉中由葉綠體基因突變或不規則分離產生的色素花斑嵌合體。②核基因嵌合體,如小鼠皮毛色斑、飛燕草玫瑰紅花斑上的紫色條紋嵌合體等。這些都是核基因突變后在正常遺傳背景上出現的變異組織的嵌合體。
嵌合體過程
整體嫁接 高等植物中取得異源嵌合體的常用方法。一般將接穗和砧木的癒合部位橫截為二,在斷面上形成的愈傷組織常能分化並長出新芽。凡是具有嫁接雙方特徵的新枝都是嵌合體。如果外層組織原層來自嫁接一方,其餘原層屬於嫁接另一方則發育為周緣嵌合體;如果全部組織原層有一個區段為一方,其餘的為另一方便發育為扇形嵌合體;如果某一層的一部分為一方,其餘為另一方,或是雙方組織在構成組織原層時相互鑲嵌,便發育為周緣區段嵌合體,如果雙方組織在空間位置上呈不規則分佈便成為花斑嵌合體。
組織併合 1972年P.S.卡爾森等將栽培煙草(Nico-tiana tabacum) 和雜種雙倍體煙草(N.glauca ×N.Langsdorfii)的髓組織緊貼培養,挑選有雙方特徵的嵌合體組織塊繼代培養,可使之分化成為嵌合體小植株。將不同基因型的愈傷組織塊緊貼培養,或以酶處理使雙方愈傷組織細胞解離后相互混合,再行培養后也能得到類似結果。
胚胎併合 1967年美國學者B.明茨從純系黑鼠和純系白鼠的輸卵管中分別取出8細胞期到桑椹期的胚胎,在試管內用蛋白酶去除透明帶,使兩個幼胚緊密接觸並融為一體,在體外繼續培養到胚泡階段,然後移植到經孕激素處理的母鼠子宮內,所獲得的皮毛黑白混生的嵌合體小鼠稱為異表型嵌合體。
移植 用射線照射使受體哺乳動物失去免疫特異性,然後將具有正常活力的另一基因型的供體骨髓細胞植入受體的骨髓中,受體內兩種基因型的漿細胞有時能夠長期存活而構成漿細胞嵌合體。

嵌合體
由兩種或多種遺傳上不同的細胞類型混合而成的植物體或植物部分。嵌合體可為「嫁接雜種」,即在植物嫁接時出現於接穗與砧木交接處的芽,包含兩種植物的組織。雖然這種嵌合體在過去偶然出現過,但是德國植物學家溫克勒(Hans Winkler)於1907年才首次認真地研究這種嵌合體。在他最早進行的一些實驗中,他將龍葵(Solanum nigrum)嫁接到番茄(Solanum lycopersicum)上,在連接處發出的枝條或具龍葵的特徵或具番茄的特徵,只有一個從兩種植物組織交界處發出的枝條,在一側具龍葵的特徵,另一側具有番茄的特徵。溫克勒稱之為嵌合體,因為它在一定程度上應屬某一個種,在一定程度上又屬另一個種。在以後進一步的實驗中,他給某些嫁接雜種起了專門名字。另一位植物學家鮑爾(Erwin Baur)後來提出證據說明︰溫克勒起了名的兩種植物分別由番茄核心和有1~2層細胞厚的龍葵外皮組成,而另外兩種則由龍葵核心和1~2層細胞厚的番茄外皮組成。這樣,在嵌合體中各個組成成分保持它們的特性,但在生長點處以一定的形式排列。
嵌合體亦可因生長區細胞的突變而形成。新的組織類型可以與舊類型明顯不同(例如舊類型為綠色而新類型為無色),但更常見的情況是︰這種差異僅能用特殊的研究手段發現,例如染色體數目的改變。
另:廣義的嵌合體(chimera)也包括在基因克隆時,由於PCR過程中的某些因素,造成的同一克隆片段來源於不同物種的現象,常見於微生物多樣性分析基因克隆時。
基本信息
- 中文名
- 嵌合體
- 外文名
- Chimera
- 定義
- 不同遺傳性狀嵌合或混雜的個體
- 應用
- 園藝 育種
- 類別
- 同源嵌合體 異源嵌合體
- 具體實例
- 單嵌合體分佈 多嵌合體分佈