光合磷酸化
光合磷酸化
光合磷酸化是指由光照引起的電子傳遞與磷酸化作用相偶聯而生成ATP的過程。
光合磷酸化(photophosphorylation)是植物葉綠體的類囊體膜或光合細菌的載色體在光下催化腺二磷(ADP)與磷酸(Pi)形成腺三磷(ATP)的反應。有兩種類型:循環式光合磷酸化和非循環式光合磷酸化。前者是在光反應的循環式電子傳遞過程中同時發生磷酸化,產生ATP。後者是在光反應的非循環式電子傳遞過程中同時發生磷酸化,產生ATP。在非循環式電子傳遞途徑中,電子最終來自於水,最後傳到氧化型輔酶Ⅱ(NADP+)。因此,在形成ATP的同時,還釋放了氧並形成還原型輔酶Ⅱ(NADPH)。
在光合作用的光反應中,除了將一部分光能轉移到NADPH中暫時儲存外,還要利用另外一部分光能合成ATP,將光合作用與ADP的磷酸化偶聯起來,這一過程稱為光合磷酸化。它同線粒體的氧化磷酸化的主要區別是:氧化磷酸化是由高能化合物分子氧化驅動的,而光合磷酸化是由光子驅動的。
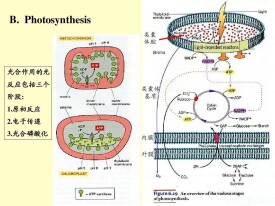
光合磷酸化的機理同線粒體進行的氧化磷酸化相似,同樣可用化學滲透學說來說明。在電子傳遞和ATP合成之間, 起偶聯作用的是膜內外之間存在的質子電化學梯度。類囊體膜進行的光合電子傳遞與光合磷酸化需要四個跨膜複合物參加:光系統Ⅱ、細胞色素b6/f複合物、光系統Ⅰ和ATP合酶。有三個可動的分子(質子):質體醌、質體藍素和H+質子將這四個複合物在功能上連成一體:即完成電子傳遞、建立質子梯度、合成ATP和NADPH。
與光合電子傳遞類同,光合磷酸化也被分為三種類型。
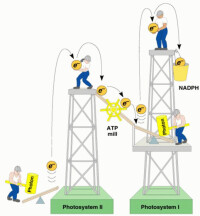
1.非環式光合磷酸化(noncyclic photophosphorylation) 與非環式電子傳遞偶聯產生ATP的反應。按圖4-15,非環式光合磷酸化與吸收量子數的關係可用下式表示。
2NADP+3ADP+3Pi+2HO → 2NADPH+2H+3ATP+O 在進行非環式光合磷酸化的反應中,體系除生成ATP外,同時還有NADPH的產生和氧的釋放。非環式光合磷酸化僅為含有基粒片層的放氧生物所特有,它在光合磷酸化中佔主要地位。
2.環式光合磷酸化(cyclic photophosphorylation) 與環式電子傳遞偶聯產生ATP的反應。
ADP+Pi → ATP+HO
環式光合磷酸化是非光合放氧生物光能轉換的唯一形式,主要在基質片層內進行。它在光合演化上較為原始,在高等植物中可能起著補充ATP不足的作用。
3.假環式光合磷酸化(pseudocyclic photophosphorylation) 與假環式電子傳遞偶聯產生ATP的反應。此種光合磷酸化既放氧又吸氧,還原的電子受體最後又被氧所氧化。
HO+ADP+Pi → ATP+ O。+4H+NADP+供應量較低,例如NADPH的氧化受阻,則有利於假環式電子傳遞的進行。
非環式光合磷酸化與假環式光合磷酸化均被DCMU(二氯苯基二甲基脲,dichlorophenyl dimethylures,商品名為敵草隆,diuron,一種除草劑)所抑制,而環式光合磷酸化則不被DCMU抑制。
1966年,Andre Jagendorf實驗證明,即使在暗處葉綠體也可以形成人ATP,只要在類囊膜兩側形成人為的pH梯度。即將葉綠體在pH4緩衝液中泡12小時,然後迅速與含ADP、Pi的pH 8緩衝液混合,葉綠體基質的pH迅速升至8,但是類囊體中的pH仍是4,這時發現隨著類囊膜兩側pH梯度的消失,同時有ATP形成,所以提出與氧化磷酸化類似的光合磷酸化的化學滲透學說。
在光合磷酸化中也需要完整的膜,在光激發下H+從基質流向類囊膜內形成跨膜質子梯度。ATP酶是在膜外,形成ATP后后質子才流出去。?
不同結構中的ATP酶
 2張
2張不同波長下光量子具有的能量不同
光合作用的總反應式為:
6CO + 6HO ——→ CHO + 6O
ΔG=2881千焦耳/摩爾
每摩爾光量子含有6.02×10^23光量子,不同波長下光量子具有的能量不同。短波長光能量較大。
若按700nm波長光計算,48個光量子有48×170=8265千焦耳/摩爾能量,在標準條件下,1摩爾葡萄糖需花2881千焦耳自由能,因此光合作用能量利用效率為288l/8265%=35%。
光合磷酸化與電子傳遞的偶聯關係
三種光合磷酸化作用都與電子傳遞相偶聯。如果在葉綠體體系中加入電子傳遞抑製劑,那麼光合磷酸化就會停止;同樣,在偶聯磷酸化時,電子傳遞則會加快,所以在體系中加入磷酸化底物會促進電子的傳遞和氧的釋放。
磷酸化和電子傳遞的關係可用ATP/e2-或P/O來表示。ATP/e2-表示每對電子通過光合電子傳遞鏈而形成的ATP分子數;P/O表示光反應中每釋放1個氧原子所能形成的ATP分子數。比值越大,表示磷酸化與電子傳遞偶聯越緊密。從圖4-15或(4-27)式看,經非環式電子傳遞時分解2分子H2O,放1 個O2與傳遞2對電子,使類囊體膜腔內增加8個H(放氧複合體處放4個H,PQH2與Cytb6/f間的電子傳遞時放4個H),如按8個H形成3個ATP算,即傳遞2對電子放1個O,能形成3個ATP,即ATP/e2或P/O理論值應為1.5,而實測值是在0.9~1.3之間。
化學滲透學說
關於光合磷酸化的機理有多種學說,如中間產物學說、變構學說、化學滲透學說等,其中被廣泛接受的是化學滲透學說。
化學滲透學說(chemiosmotic theory)由英國的米切爾(Mitchell,1961)提出,該學說假設能量轉換和偶聯機構具有以下特點:①由磷脂和蛋白多肽構成的膜對離子和質子的透過具有選擇性 ②具有氧化還原電位的電子傳遞體不勻稱地嵌合在膜內。③膜上有偶聯電子傳遞的質子轉移系統。④膜上有轉移質子的ATP酶。在解釋光合磷酸化機理時,該學說強調:光合電子傳遞鏈的電子傳遞會伴隨膜內外兩側產生質子動力(proton motive force,pmf),並由質子動力推動ATP的合成。許多實驗都證實了這一學說的正確性。
(1)化學滲透學說的實驗證據
①階段光合磷酸化實驗
指光合磷酸化可以相對分成照光階段和暗階段來進行,照光不向葉綠體懸浮液中加磷酸化底物,而斷光時再加入底物能形成ATP的實驗。1962年,中國的沈允鋼等人,用此實驗探測到光合磷酸化高能態(Z*)的存在。1963年賈格道夫(Jagendorf)等也觀察到了光合磷酸化高能態的存在。起初認為Z*是一種化學物質,以此提出了光合磷酸化中間物學說。現在知道高能態即為膜內外的H+電化學勢。所謂兩階段光合磷酸化,其實質是光下類囊體膜上進行電子傳遞產生了跨膜的H+電化學勢,暗中利用H+電化學勢將加入的ADP與Pi合成ATP。
②酸-鹼磷酸化實驗
賈格道夫等(1963)在暗中把葉綠體的類囊體放在pH4的弱酸性溶液中平衡,讓類囊體膜腔的pH下降至4(圖4-16A),然後加進pH8和含有ADP和Pi的緩衝溶液(圖4-16B),這樣瞬間的pH變化使得類囊體膜內外之間產生一個H+梯度。這個H+梯度能使ADP與Pi生成ATP,而這時並不照光,也沒有電子傳遞。這種驅動ATP合成的類囊體內外的pH差在活體中正是由光合電子傳遞和H+轉運所形成的。這一酸-鹼磷酸化實驗給化學滲透假說以最重要的支持證據。
③光下類囊體吸收質子的實驗
對無pH緩衝液的葉綠體懸浮液照光,用pH計可測到懸浮液的pH升高。這是由於光合電子傳遞引起了懸浮液中質子向類囊體膜腔運輸,使得膜內H+濃度高而膜外較低的緣故。電子傳遞產生了質子梯度后,質子就有反向跨膜轉移的趨向,質子反向轉移時,質子梯度所貯藏的能量就被用去合成ATP(圖4-15)。
以上實驗都證實了米切爾的化學滲透學說的正確性,因而米切爾獲得了1978年度的諾貝爾化學獎。
(2)H+電化學勢與質子動力
ΔμH+=μH+內 -μH+外
=RT(ln [H+內] -ln [H+外])+F(E內-E外)
=RTln([H+內]/ [H+外])+FΔE
=2.3RTΔpH+FΔE (4-30)
式中R-氣體常數(8.314J·mol-1·K-1),T-絕對溫度(K),F-法拉第常數(96.5kJ·mol-1·V-1),ΔE-膜電勢(V)。
25℃時,ΔμH+ =5.7ΔpH(kJ·mol-1)+96.5ΔE(kJ·mol-1) (4-31)
上式(4-31)中5.7ΔpH(kJ·mol-1)為膜內外質子濃度差所具有的能量,而96.5ΔE(kJ·mol-1)為膜電勢所具有的能量。
將式4-31兩邊用F(96.5kJ·mol-1·V-1)除,規定△μH+/F為質子動力,其單位為電勢(V)。
在25℃時:pmf=0.059ΔpH+ΔE
葉綠體類囊體膜的質子動力大部分是來自ΔpH部分,電荷分佈所產生的ΔE的貢獻很小,原因是其它離子,如Cl-、K+或Mg2+也能穿透類囊體膜,當H+穿透類囊體膜時,Cl-可以與H+同向穿透,或Mg2+ 與H+(1Mg2+/2H+)反向穿透,這樣就保持了電中性,結果不產生電勢差。
ATP合成的部位——ATP酶
質子反向轉移和合成ATP是在ATP酶(腺苷三磷酸酶 adenosine triphosphatase,ATPase)上進行的。葉綠體內囊體膜上的ATP酶也稱偶聯因子(coupling factor)或CF1-CF0複合體。葉綠體的ATP酶與線粒體、細菌膜上的ATP酶結構十分相似,都由兩個蛋白複合體組成:一個是突出於膜表面的親水性的“CF1”;另一個是埋置於膜中的疏水性的“CF0”。ATP酶由九種亞基組成,分子量為550 000左右,催化的反應為磷酸酐鍵的形成,即把ADP和Pi合成ATP。另外ATP酶還可以催化逆反應,即水解ATP,並偶聯H+向類囊體膜內運輸。
CF1的分子量約400 000,它含有α(60 000),β(56 000),γ(39 000),δ(19 000)和ε(14 000)的5種亞基。其中α亞基有結合核苷酸的部位,在進行催化時可能發生構象變化;β亞基是合成和水解ATP分子的催化位置;γ亞基控制質子的穿流;δ亞基也許與CF0的結合有關;ε亞基似乎能抑制CF1-CF0複合體在暗中的活性,防止ATP的水解。δ和ε亞基還有阻塞經CF0的質子泄漏的作用。 CF0含有四個亞基:Ⅰ、Ⅱ、Ⅲ和Ⅳ。Ⅲ是多聚體,可能含有12個多肽,總分子量為100 000。Ⅲ可能是CF0中質子轉移的主要通道,而Ⅰ、Ⅱ、Ⅳ亞基的功能可能與建立質子轉移通道或與結合CF1有關。
當類囊體膜失去CF1后,就失去磷酸化功能,如果重新加進CF1即可恢復磷酸化功能。失去了CF1的類囊體膜會泄漏質子。但是一旦將CF1加回到膜上或是加進CF0的抑製劑后,質子泄漏就停止了。這表明CF0是質子的“通道”,供應質子給CF1去合成ATP。至於CF1如何利用H+越膜所釋放的能量來合成ATP,美國的鮑易爾(Boyer 1993)認為,是H+濃度遞度引起CF1上亞基的轉動變構而催化ATP合成的。
ATP合成的結合轉化機制
γ-亞基的轉動引起β亞基的構象依緊繃(T)、鬆弛(L)和開放(O)的順序變化,完成ADP和Pi的結合、 ATP的 形成以及ATP的釋放三個過程
光合磷酸化的抑製劑
葉綠體進行光合磷酸化,必須:(1)類囊體膜上進行電子傳遞;(2)類囊體膜內外有質子梯度;(3)有活性的ATP酶。破壞這三個條件之一的試劑都能使光合磷酸化中止,這些試劑也就成了光合磷酸化的抑製劑。
(1)電子傳遞鏈
傳遞過程是:P680→pheo→Q→PQ→Fe-S-Cytb6→Cytf→PC→P700。其中 P700 和 P680
分別為光系統Ⅰ和光系統Ⅱ的作用中心色素,pheo為去鎂葉綠素
(2)電子傳遞抑製劑
指抑制光合電子傳遞的試劑,如羥胺(NH2OH)切斷水到PSⅡ的電子流,DCMU抑制從PSⅡ上的Q到PQ的電子傳遞;KCN和Hg等則抑制PC的氧化。一些除草劑如西瑪津(simazine)、阿特拉津(atrazine)、除草定(bromacil)、異草定(isocil)等也是電子傳遞抑製劑,它們通過阻斷電子傳遞抑制光合作用來殺死植物。
(3)解偶聯劑
指解除磷酸化反應與電子傳遞之間偶聯的試劑。常見的這類試劑有DNP(dinitrophenol,二硝基酚)、CCCP(carbonyl cyanide-3-chlorophenyl hydrazone,羰基氰-3-氯苯腙)、短桿菌肽D、奈及利亞菌素、NH等,這些試劑可以增加類囊體膜對質子的透性或增加偶聯因子滲漏質子的能力,其結果是消除了跨膜的H+電化學勢,而電子傳遞仍可進行,甚至速度更快(因為消除了內部高H+濃度對電子傳遞的抑制),但磷酸化作用不再進行。
(4)能量傳遞抑製劑
指直接作用ATP酶抑制磷酸化作用的試劑,如二環己基碳二亞胺(DCCD)、對氯汞基苯(PCMB)作用於CF1,寡黴素作用於CF(CF 下標的o就是表明其對寡黴素oligomycin敏感)。它們都抑制了ATP酶活性從而阻斷光合磷酸化。
葉綠體電子傳遞鏈的抑製劑作用位點
基本信息
- 中文名
- 光合磷酸化
- 外文名
- Photophosphorylation
- 目的
- 生成ATP的過程
- 兩種類型
- 循環和非循環式光合磷酸化