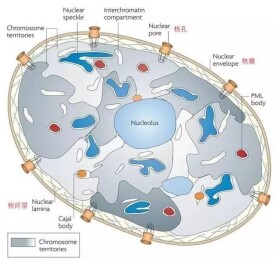
核纖層
細胞核內層核膜下的纖維網路
核纖層由核纖肽(lamin)構成,核纖肽分子量約60~80KD,是一類中間纖維,在哺乳類和鳥類中可分為A、B兩型。核纖層的作用有以下兩個方面:保持核的形態和參與染色質和核的組裝。
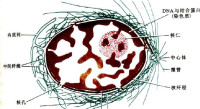
核纖層
間期細胞核中,核纖層與核膜在結構上有密切聯繫,常與核膜組分一起被分離。有人認為廣義的核膜包括3種主要結構組分:(1)核外膜和核內膜;(2)核孔複合體;(3)核纖層,而後兩者又被認為是核骨架的有機組分。當用高鹽、非離子去垢劑和核酸酶處理核膜后,只有核膜孔複合體和核纖層存留,核纖層對鹽溶液有較大的穩定性,在細胞抽提中常與中間纖維、核骨架共同被分離。
核纖層的研究始於50年代,而真正取得實質性進展是在70年代末和80年代。lamina曾有許多不同的名稱,如fibrous lamina、dense lamella、zona nuclear limitans,目前通用的名稱是lamina或nuclearlamina。現在還沒有統一的合適的中文譯名,有“核纖層”、“核層”、“核襯層”與“核周層”等不同的譯名,傾向於用核纖層來表示。
1.保持核的形態:是核被膜的支架,用高鹽溶液、非離子去污劑和核酸酶去除大部分核物質,剩餘的核纖層仍能維持核的輪廓。此外,核纖層與核骨架以及穿過核被膜的中間纖維相連,使胞質骨架和核骨架形成一連續網路結構。
2.參與染色質和核的組裝:核纖層在細胞分裂時呈現出周期性的變化,在間期核中,核纖層提供了染色質(異染色質)在核周邊錨定的位點。在前期結束時,核纖層被磷酸化,核膜解體。其中B型核纖肽與核膜殘餘小泡結合,A型溶於胞質中。在分裂末期,核纖肽去磷酸化重新組裝,介導了核膜的重建。
在某些真核細胞中,通過超薄切片電鏡觀察可以直接觀察到位於內層核膜與染色質之間的核纖層結構,厚度為30~100nm,然而,絕大多數細胞的核纖層是很薄的結構。只有將核膜與染色質去除后才能觀察到。免疫細胞化學研究說明,核纖層至少在高等真核細胞間期細胞核中是普遍存在的。
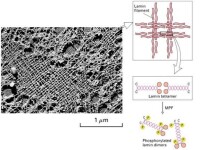
核纖層纖維的直徑為10nm左右,縱橫排列整齊,呈正交狀編織成網路,分佈於內層核膜與染色質之間。一般認為核纖層結構整體觀呈一球狀或籠狀網路,切面觀呈片層結構。在分裂期細胞,核纖層解體,以單體形式存在於胞質中。
核纖層
核纖層蛋白的分子結構及其與中間纖維蛋白的關係
早在60年代就知道核纖層具有纖維網狀結構,但對其分子基礎並不清楚。近年來發現核纖層與中間纖維有許多共同點:
(1)兩者均形成10nm纖維;
(2)兩者均能抵抗高鹽和非離子去垢劑的抽提;
(3)某些抗中間纖維蛋白的抗體能與核纖層發生交叉反應,說明中間纖維蛋白與核纖層蛋白分子存在相同的抗原決定簇(Osborn& Weber,1987);
(4)兩者在結構上有密切聯繫,核纖層成為核骨架與中間纖維之間的橋樑。
核纖層蛋白在細胞分化中的表達
核纖層
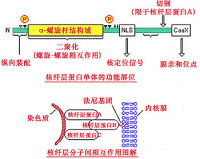
核纖層最顯著的結構重組發生於分裂期,在細胞分裂過程中,核纖層發生解聚和重裝配。分裂前期,核膜崩解,核纖層解聚,核纖層蛋白彌散到胞質中。分裂末期,當核膜重現時,在染色體周圍重裝配,形成於細胞的核纖層。
細胞分裂期中,核纖層蛋白的磷酸化水平發生顯著改變,分裂前期高度磷酸化,而末期則發生去磷酸化,提示磷酸化可能是分裂期中核纖層結構動態變化的調控因素,而核纖層蛋白磷酸化可能是分裂前期細胞內磷酸化級聯放大反應的一個重要環節,這種磷酸化級聯反應可以在時間上協調許多細胞結構在分裂期的重組。最新的研究表明,核纖層蛋白是促分裂因子(MPF)P34cdc2亞基的直接作用底物,P34cdc2亞基具蛋白激酶活性,有絲分裂前期,MPF使核纖層蛋白分子桿狀區兩端的Ser22和Ser392磷酸化,直接導致核纖層解聚。
核纖層的磷酸化與核被膜解體
核纖層
脊椎動物細胞中有三種類型的核纖層蛋白(A,B,C),核纖層蛋白。
A和C是由同一個轉錄單位編碼的,只不過是通過可變剪接形成不同的mRNA。它們在肽鏈上的差別是:核纖層蛋白A的C末端比核纖層蛋白C的C末端多133個氨基酸殘基。核纖層蛋白B是由另一個轉錄單位編碼的,通過轉錄后的修飾,在羧基端添加了疏水的異丙基,添加的脂肪酸幫助核纖層蛋白B插入到核膜的內脂層。三種類型的核纖層蛋白都以二聚體的形式存在,有球形的頭和尾部結構域以及一個桿狀的α螺旋中心。這些核纖層蛋白二聚體以頭-頭、尾-尾相接的方式形成核纖層。
脊椎動物細胞中有三種類型的核纖層蛋白(A,B,C),核纖層蛋白A和C是由同一個轉錄單位編碼的,通過可變剪接形成不同的mRNA。
核纖層的磷酸化
在有絲分裂早期,MPF使三種核纖層蛋白的特定絲氨酸殘基磷酸化,引起核纖層的去聚合磷酸化的核纖層蛋白A和C被釋放出來進入溶液中,而磷酸化的核纖層蛋白B由於異丙基的作用仍然與核膜相連。核纖層的解體,導致核被膜破裂形成小囊泡。
核纖層與核膜、染色質及核孔複合體在結構上有密切聯繫,一般認為核纖層在細胞核中起支架作用,為核膜及染色質提供了結構支架。
核纖層的作用:
1.結構支撐功能:核纖層蛋白形成骨架結構支撐於核被膜的內側,使得核被膜能起到細胞核與細胞質之間的隔離與信息交換功能。
2.調節基因表達:果蠅細胞中基因組範圍的研究結果表明,沉默基因更傾向於分佈於核纖層附近,異染色質更易與核纖層結合,而且核纖層附近的染色質的乙醯化水平較低。然而,在酵母細胞中活躍轉錄的基因也分佈於核纖層附近,它們常與核孔複合體結合。所以,核纖層與基因表達的確切關係還不是非常清楚。可能在不同物種的細胞中,甚至不同組織的細胞中情況不一樣。
4.與細胞周期的關係:細胞分裂過程中,核纖層蛋白解聚成可溶的單體或與崩解后的核被膜相結合。新核形成時,核被膜與染色質結合的同時,核纖層也最後形成。
核纖層與核膜在結構和功能上的關係是一個令人感興趣的問題。生化分析發現核纖層蛋白A,B和C均有親膜結合作用,而以核纖層蛋白B與膜的結合能力最強。在細胞分裂期中,核膜與核纖層的行為密切相關,核膜崩解和核纖層解聚后,核纖層蛋白B能選擇性地與核膜小泡結合。Gerace等(1986)推測分裂期中核纖層的可逆性解聚與重裝配對核膜的崩解與重裝配有調節作用,分裂前期核纖層的解聚為核膜崩解形成膜泡提供了必要的微環境。分裂期中核纖層蛋白B結合在核膜小泡上。分裂末期,當核纖層蛋白去磷酸化與重裝配時,直接介導了核膜圍繞染色體的重裝配,核纖層蛋白將核膜小泡“引導”到染色體表面,經膜融合后構成新核膜。這樣一個以核纖層為基礎的核膜重建模型認為少數幾種核膜相關蛋白即足以使核膜在分裂期重建。當然,這一模型尚缺乏充分的實驗證據。
A型核纖層蛋白前體
2010年5月12日英國研究人員日前報告說,他們發現一種使人衰老的蛋白質在心臟病發病過程中起著重要作用,這將有助於研發治療心臟病的新途徑。
英國倫敦國王學院等機構研究人員在新一期美國《循環》醫學期刊上報告說,以前研究曾發現名為“A型核纖層蛋白前體”的蛋白質與人體衰老有關,本次研究分析了這種蛋白質對心臟細胞的作用,發現它會使得心臟細胞難以修復自身損傷,導致心血管硬化和脂肪堆積,最終導致心臟病。
研究人員表示,這項研究為治療心臟病找到了一個新途徑,即可以通過有效抗氧化使用較強的抗氧化劑(如類胡蘿蔔素的β-胡蘿蔔素、ASTA蝦青素,番茄紅素等)減少這種蛋白質或抑制其發揮作用來延緩心臟細胞衰老。
研究還發現,在老人的心臟平滑肌細胞中往往可以發現這種蛋白質的含量偏高,這也許是老人成為心臟病高發群體的原因之一。
領導研究的凱瑟琳·沙納漢教授說,雖然這種蛋白質在體內積累與自然衰老密切相關,但人們的生活方式也會對這種蛋白質的積累速度造成影響,如吸煙等不良生活習慣就會加速它的積累,提前造成心臟細胞衰老並導致心臟病。
普遍存在於高等真核細胞中,是內層核被膜下纖維蛋白片層,其纖維直徑為10毫微米左右,纖維縱橫排列整齊呈纖維網路狀。核纖層在核內與核基質連接,在核外與中間纖維相連,構成貫穿於細胞核和細胞質的統一網架結構體系。它位於內層核膜與染色質之間,與核膜、染色質及核孔複合體在結構上有密切聯繫,核纖層蛋白向外與內層核膜上的蛋白結合,向內與染色質的特定區段結合。其厚度隨不同細胞而異,為30~100毫微米。大多數真核細胞的核纖層很薄。高等動物核纖層通常由3種屬於中間纖維的多肽組成,即核纖層蛋白(lamins)A、B、C。分子量60000~80000道爾頓。核纖層與核被膜的穩定、維持核孔位置、穩定間期染色質形態與空間結構、染色質構建和細胞核組裝密切相關。如在間期細胞中,核纖層為核膜提供了支架的作用,核纖層的可逆性解聚調節了核膜的崩解和重建,當細胞進行有絲分裂時,核纖層蛋白被磷酸化,引起核纖層可逆解聚,核膜崩解,在分裂末期時,核纖層蛋白去磷酸化,它直接介導了核膜圍繞染色體之重建。
核纖層蛋白在人的各種組織和細胞中已有比較系統的研究。其中,核纖層蛋白A/C在大多數分化良好的上皮細胞中表達,而在神經內分泌起源的大多數細胞及造血細胞中不表達。核纖層蛋白B1在肌肉和連接組織中不表達,在上皮細胞中僅表達於增殖的細胞。除了肝細胞外,核纖層蛋白B2幾乎在所有檢測的組織中表達。核纖層蛋白在各種組織中特異性的表達也許預示著其功能的特異性。
在哺乳動物中,核纖層蛋白家族包括A,A△10,C,C2,B1,B2及B3。在小鼠精子發生的過程中,各種核纖層蛋白的表達有特異性。其中,核纖層蛋白B2在整個精子發生過程中表達,但是核纖層蛋白A、C、B2不表達。核纖層蛋白C2僅特異性地表達於減數分裂期,而核纖層蛋白B3僅在精子發生後期即精子形成期表達。在睾丸的曲細精管中,支持細胞、精原細胞及各級減數分裂期精母細胞不表達核纖層蛋白B3,在圓形精子中可以檢測到核纖層蛋白B3。免疫熒光結果表明,核纖層蛋白B3的熒光信號分佈於細胞核的邊緣,並且在著絲粒異染色質上也有微弱的信號。隨著精子細胞的進一步成熟,熒光信號逐漸移向圓形精子和延長形精子的後部。核纖層蛋白B3位置改變的生物學功能還有待於進一步研究。
有研究表明,A型核纖層蛋白是雄性小鼠生殖能力的重要決定因素。與野生型及lmna+/-雜合型小鼠相比,lmna-/-小鼠的睾丸體積及曲細精管的直徑明顯減小。雖然精原細胞和支持細胞數量基本未發生改變,但是I型精母細胞的數量減少。此外,減數分裂後期的細胞(精子細胞和精子)完全消失。TUNEL實驗結果表明,lmna-/-小鼠曲細精管中凋亡細胞的數量增多。雖然lmna-/-小鼠附睾中也充滿了官腔液,但是液體中不含精子,而在LMNA基因缺失的雌性小鼠中,卵子發生正常。
維持細胞核形狀
許多研究表明,核纖層對於細胞核形狀的維持是至關重要的。在小鼠精母細胞中,精母細胞特異性表達的核纖層蛋白B3缺失后導致細胞核形狀從球形變為鉤形。實際上,在COP5成纖維細胞中表達外源的核纖層蛋白B3也可以導致細胞核形態發生變化。在編碼核纖層蛋白A/C基因敲除的小鼠和秀麗新小桿線蟲中,同樣可以觀察到細胞核形態的改變。此外,人HIV病毒VPR蛋白可以誘導核纖層局部解體,最終導致核孔上出現可見的泡狀物。
維持細胞核大小
在非洲爪蟾卵細胞核提取物中,核纖層蛋白控制細胞核的大小。在細胞核提取物中核纖層被免疫缺失后,雖然核膜的組裝仍然進行,但是細胞核變小。
參與核膜的組織
超微結構研究發現,核纖層纖絲在二維空間上互相垂直排列,交織成網格狀結構,並且與核孔複合物(NPC)的某些亞基相互作用,從而把NPC錨定在核膜的相應位置上。在果蠅中,核纖層蛋白Dm0缺失后,可以觀察到NPC在核膜上漂浮,並最終聚集到一起。同樣,在lmn-1缺失的線蟲中也觀察到類似的結果。
把核纖層蛋白徵集到核纖層上
許多研究表明,在內層核膜中,核纖層蛋白之間的相互作用也有先後次序。其中,B類核纖層蛋白首先結合到核纖層纖絲上,然後是核纖層蛋白A,最後是核纖層蛋白C。在非洲爪蟾的卵細胞提取物中,重組的核纖層蛋白A僅當內源性核纖層蛋白B3存在的情況下,才能結合到體外組裝的精子原核的核膜上,這表明核纖層蛋白A也許僅能結合到已經存在的B型核纖層蛋白纖絲上。另外,還有證據表明,在體外培養的細胞中,核纖層蛋白A在B型核纖層蛋白組裝到有絲分裂末期的細胞核前,一直定位於核質中。核纖層蛋白A和C的組裝特性也不同。在小鼠細胞中表達或注射標記的核纖層蛋白后可以發現,核纖層蛋白A比核纖層蛋白C能夠更快的組裝到核纖層上。這些研究表明,核纖層蛋白A可以指導核纖層蛋白C的組裝。
參與DNA的複製
在非洲爪蟾卵細胞提取物中,核纖層蛋白B3的缺失將會導致細胞核體積縮小,並且生物素標記的dUTP也不能夠積極地參與到DNA複製過程中。當把純化的核纖層蛋白B3重新加入提取物中,細胞核形狀逐漸恢復,並且DNA複製重新開始。在體細胞有絲分裂的S期,核纖層蛋白B1從核膜轉移到DNA複製中心。這些結果表明,核纖層蛋白在DNA複製過程也許發揮了直接的作用。
參與有絲分裂
核纖層蛋白是促分裂因子(MPF)P34cdc2亞基的直接作用底物,P34cdc2亞基具蛋白激酶活性,有絲分裂前期,MPF使核纖層蛋白分子桿狀區兩端的Ser22和Ser392磷酸化,直接導致核纖層解聚及核膜崩解。在分裂末期時,核纖層蛋白去磷酸化,它直接介導了核膜圍繞染色體之重建。
細胞結構與細胞外基質
| ▪ 超微結構 | ▪ 顯微結構 | ▪ [細]胞間橋 | ▪ 生物膜 | ▪ 單位膜 |
| ▪ 細胞膜 | ▪ 質膜 | ▪ 胞質面 | ▪ 質膜外面 | ▪ 表膜 |
| ▪ 間體 | ▪ 脂單層 | ▪ 脂雙層 | ▪ 脂筏 | ▪ 膜流動性 |
| ▪ 磷脂雙層 | ▪ 膜再循環 | ▪ 細胞外被 | ▪ 微絨毛 | ▪ 刷狀緣 |
| ▪ 菌毛 | ▪ 纖毛 | ▪ 靜纖毛 | ▪ 根絲體 | ▪ 纖毛小根系統 |
| ▪ 鞭毛 | ▪ 軸絲 | ▪ 基體 | ▪ 副基體 | ▪ 動基體 |
| ▪ 基片 | ▪ km纖維 | ▪ 偽足 | ▪ 絲足 | ▪ 微棘 |
| ▪ 葉足 | ▪ 片足 | ▪ 足體 | ▪ 軸足 | ▪ 胞口 |
其他科技名詞
| ▪ 胞咽 | ▪ 胞肛 | ▪ 陷窩 | ▪ 陷窩蛋白 | ▪ 細胞連接 |
| ▪ 緊密連接 | ▪ 黏著連接 | ▪ 橋粒 | ▪ 半橋粒 | ▪ 黏著斑 |
| ▪ 黏著帶 | ▪ 張力絲 | ▪ 間隙連接 | ▪ 連接子 | ▪ 分隔連接 |
| ▪ 細胞黏附 | ▪ 細胞黏附分子 | ▪ 外質體 | ▪ 原生質體 | ▪ 亞原生質體 |
| ▪ 共質體 | ▪ 合胞體 | ▪ 質外體 | ▪ 乳突 | ▪ 毛狀體 |
| ▪ 微原纖維 | ▪ 細胞壁 | ▪ 初生細胞壁 | ▪ 次生細胞壁 | ▪ 珠光壁 |
| ▪ 紋孔場 | ▪ 篩域 | ▪ 篩板 | ▪ 篩孔 | ▪ 胞壁內突生長 |
| ▪ 凱氏帶 | ▪ 花粉外壁 | ▪ 花粉內壁 | ▪ 孢子中壁 | ▪ 孢子外壁 |
| ▪ 萌發孔 | ▪ 吸器 | ▪ 胞間連絲 | ▪ 初生胞間連絲 | ▪ 次生胞間連絲 |
| ▪ 連絲小管 | ▪ [胞]外連絲 | ▪ 烏氏體 | ▪ 胞質孔環 | ▪ 頸區 |
| ▪ 共質域 | ▪ 外質 | ▪ 內質 | ▪ 原生質 | ▪ 細胞質 |
| ▪ 皮質 | ▪ 皮層 | ▪ 周質間隙 | ▪ 周質 | ▪ 胞質溶膠 |
| ▪ 細胞器 | ▪ 線粒體 | ▪ 線粒體內膜 | ▪ 線粒體基質 | ▪ 線粒體外膜 |
| ▪ 線粒體嵴 | ▪ 亞線粒體小泡 | ▪ 線粒體核糖體 | ▪ 肌粒 | ▪ OXA複合體 |
| ▪ TOM複合體 | ▪ TIM複合體 | ▪ 膜間隙 | ▪ 葉綠體 | ▪ 葉綠體被膜 |
| ▪ 葉綠體基質 | ▪ 葉綠體基粒 | ▪ 基質片層 | ▪ 基粒片層 | ▪ 類囊體 |
| ▪ 基粒類囊體 | ▪ 基質類囊體 | ▪ 前質體 | ▪ 質體 | ▪ 色質體 |
| ▪ 白色體 | ▪ 造粉體 | ▪ 黃化質體 | ▪ 油質體 | ▪ 圓球體 |
| ▪ 裸質體 | ▪ 糊粉粒 | ▪ 澱粉核 | ▪ 葉褐體 | ▪ 藻膽[蛋白]體 |
| ▪ 藻紅體 | ▪ 內膜系統 | ▪ 內質網 | ▪ 光面內質網 | ▪ 糙面內質網 |
| ▪ 動質 | ▪ 微粒體 | ▪ 轉運體 | ▪ Sec61複合體 | ▪ 肌質網 |
| ▪ 核糖[核蛋白]體 | ▪ 多核糖體 | ▪ 高爾基[複合]體 | ▪ 分散[型]高爾基體 | ▪ 順面 |
| ▪ 反面 | ▪ 順面高爾基網 | ▪ 反面高爾基網 | ▪ 瀦泡 | ▪ 液泡 |
| ▪ 液泡形成體 | ▪ 小泡 | ▪ 收縮泡 | ▪ 溶酶體 | ▪ 初級溶酶體 |
| ▪ 次級溶酶體 | ▪ 自噬溶酶體 | ▪ 異噬溶酶體 | ▪ 吞噬溶酶體 | ▪ 殘餘體 |
| ▪ 吞噬體 | ▪ 異[吞]噬體 | ▪ 自[體吞]噬體 | ▪ 內[吞]體 | ▪ 早期內體 |
| ▪ 晚期內體 | ▪ 再循環內體 | ▪ 多泡體 | ▪ 外排體 | ▪ 溶酶體貯積症 |
| ▪ 微體 | ▪ 過氧化物酶體 | ▪ 微過氧化物酶體 | ▪ 乙醛酸循環體 | ▪ 細胞骨架 |
| ▪ 微梁網 | ▪ 微管 | ▪ 微管組織中心 | ▪ 自組裝 | ▪ 正端 |
| ▪ 負端 | ▪ 踏車現象 | ▪ 中心體 | ▪ 中心粒 | ▪ 原中心粒 |
| ▪ 中心球 | ▪ 中心粒周物質 | ▪ 原絲 | ▪ 絲 | ▪ 動纖絲 |
| ▪ 肌線 | ▪ 微絲 | ▪ 中間絲 | ▪ 角蛋白絲 | ▪ 結蛋白絲 |
| ▪ 神經膠質絲 | ▪ 核纖層蛋白絲 | ▪ 波形蛋白絲 | ▪ [念]珠狀纖絲 | ▪ 肌絲 |
| ▪ 粗肌絲 | ▪ 細肌絲 | ▪ 肌原纖維 | ▪ 肌節 | ▪ 肌質 |
| ▪ 肌小管 | ▪ 胞內小管 | ▪ 橫小管 | ▪ 肌膜 | ▪ 明帶 |
| ▪ 暗帶 | ▪ M 線 | ▪ Z盤 | ▪ 應力纖維 | ▪ 附著斑 |
| ▪ 神經元 | ▪ 神經絲 | ▪ 核周體 | ▪ 軸突 | ▪ 樹突 |
| ▪ 突觸 | ▪ 運動終板 | ▪ 髓鞘 | ▪ 神經原纖維 | ▪ 雙核體 |
| ▪ 多形核 | ▪ [細]胞核 | ▪ 真核 | ▪ 擬核 | ▪ [配子]原核 |
| ▪ 大核 | ▪ 小核 | ▪ 微核 | ▪ 營養核 | ▪ 生殖核 |
| ▪ 管核 | ▪ 核液 | ▪ 核質 | ▪ 核質比 | ▪ 外切體 |
| ▪ 核被膜 | ▪ 外核膜 | ▪ 內核膜 | ▪ 核周隙 | ▪ 環孔片層 |
| ▪ 核孔[複合體] | ▪ [核孔複合體]中央顆粒 | ▪ 胞質環 | ▪ 核質環 | ▪ 核籃 |
| ▪ 柱狀亞單位 | ▪ 環狀亞單位 | ▪ 腔內亞單位 | ▪ 孔膜區 | ▪ 核仁 |
| ▪ 顆粒組分 | ▪ [核仁]顆粒區 | ▪ [核仁]纖維區 | ▪ [核仁]纖維中心 | ▪ [核仁]無定形區 |
| ▪ 緻密纖維組分 | ▪ 核仁結合染色質 | ▪ 核仁染色質 | ▪ 核仁組織區 | ▪ 核仁組織染色體 |
| ▪ 核球 | ▪ 核仁線 | ▪ 核仁內粒 | ▪ 核網 | ▪ 染色質 |
| ▪ 常染色質 | ▪ 異染色質 | ▪ 組成性異染色質 | ▪ 兼性異染色質 | ▪ 螺線管 |
| ▪ 核小體 | ▪ 染色質纖維 | ▪ 染色質凝縮 | ▪ 核基質 | ▪ 核纖層 |
| ▪ 染色體支架 | ▪ [細]胞外基質 | ▪ [細]胞外基質受體 | ▪ 間充質 | ▪ 結締組織 |
| ▪ 膠原 | ▪ 膠原纖維 | ▪ 膠原原纖維 | ▪ 前膠原 | ▪ 原膠原 |
| ▪ 彈性纖維 | ▪ 彈性蛋白 | ▪ 原彈性蛋白 | ▪ 蛋白聚糖 | ▪ 黏結蛋白聚糖 |
| ▪ 糖胺聚糖 | ▪ 透明質酸 | ▪ 透明質酸黏素 | ▪ 肝素 | ▪ 硫酸乙醯肝素 |
| ▪ 硫酸角質素 | ▪ 硫酸軟骨素 | ▪ 硫酸皮膚素 | ▪ 基[底]膜 | ▪ 黏附受體 |
| ▪ 纖維素 | ▪ 半纖維素 | ▪ 木聚糖 | ▪ 磷壁酸 | ▪ 糖醛酸磷壁酸 |
| ▪ 阿拉伯聚糖 | ▪ 半乳聚糖 | ▪ 阿拉伯半乳聚糖 | ▪ 果膠 | ▪ 伸展蛋白 |
| ▪ 木栓質 | ▪ 木質素 | ▪ 愈傷葡萄糖 | ▪ 基質金屬蛋白酶 | ▪ 組織金屬蛋白酶抑制物 |
基本信息
- 中文名
- 核纖層
- 外文名
- nuclear lamina
- 厚度
- 30到100毫微米
- 存在於
- 高等真核細胞中
- 直徑
- 10納米左右
- 成分
- 核纖層蛋白
- 位置
- 細胞核內層核膜下的纖維蛋白片層或纖維網路