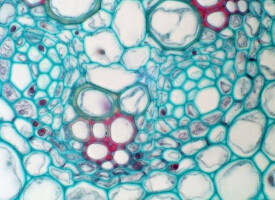
薄壁細胞
有很薄的初生壁無次生壁的細胞
薄壁細胞是多數植物體內數目最多的細胞,成熟後繼續存活,只有很薄的初生壁,沒有次生壁,一般為直徑近乎相等的多面體,但也可以分化為星芒、分枝以及臂狀等。薄壁細胞的功能很多,如貯存營養物質、進行光合作用和需氧呼吸等。
薄壁組織,薄壁細胞,葉綠體,細胞間隙,分生組織
構成植物薄壁組織的細胞,為大致等粗或多少細長的多面體,平均有14面。在幼嫩時幾乎沒有細胞間隙,以後隨著成長而逐漸形成。全為具有原生質的活細胞,細胞壁只是很薄的初生壁,具有大形液泡,進行合成、分解、貯藏等重要的生理作用。含有葉綠體的葉肉的同化組織、貯藏組織和分泌組織均由薄壁細胞組成,根據所含物質的不同,有結晶細胞、單寧細胞、乳汁細胞、油脂細胞及粘液細胞等。一般薄壁細胞來源於基本分生組織,但附屬於初生維管束和次生維管束的薄壁細胞則分別來源於原始形成層和形成層。此外,也有由木栓形成層形成的。薄壁組織細胞具有潛在的細胞分裂能力,而且在細胞間多具發達的細胞間隙,在一定的外界因素刺激下,細胞能發生反分化,恢復分生能力,轉變為分生組織(Meristem),促使植物的創傷癒合、再生,形成不定根或不定芽。在離體培養的條件下,分離的薄壁組織細胞團,甚至單個細胞,能經培養而長成一棵完整的植株。
依據薄壁組織生理機能的不同,可分為以下七種類型:
基本薄壁組織
基本薄壁組織多分佈在根、莖等器官的內部,如皮層和髓等處的起填充作用的薄壁組織。基本薄壁組織的細胞無色,橫切面呈圓球或多角狀,長與寬的差異不明顯,幾乎等徑,胞內具生活的原生質體,是營養性的生活細胞。在植物體中,基本薄壁組織起填充的作用,因而也稱其為填充薄壁組織。
同化薄壁組織
同化薄壁組織因與光合作用關係密切而得名,多存在於植物體表的易受光部位。其特點是細胞內含有葉綠體,能進行光合作用,所以又稱綠色薄壁組織(chlorenchyma)。例如植物葉上、下表皮之間的葉肉中都含有葉綠體,尤其是靠近上表皮的柵欄組織細胞中,葉綠體更多。葉綠體也存在於皮層中或莖的較內部甚至髓部。在植物幼嫩的部位如綠色的莖、枝條和果實的外部也常具一些同化薄壁組織。在個別具退化葉的植物(如麻黃、天冬)的莖中,其表皮下有數層含葉綠體的薄壁組織細胞代替葉子的同化薄壁組織細胞,常出現明顯的液泡化並形成高度的腔室結構,從而有利於進行氣體交換。
貯藏薄壁組織
貯藏薄壁組織是積存植物特殊后含物,如澱粉粒、蛋白質顆粒、擬晶體、脂肪球、油滴以及其他有機物質等的一種組織。主要分佈在根、根莖、種子和果實等器官中。甘薯的塊根、馬鈴薯的塊莖、豆類種子的子葉及穀類作物籽粒的胚乳中貯藏薄壁組織尤為發達。上述后含物一般以溶解狀態存在於貯藏薄壁組織細胞的細胞液內,有的以液體或固體狀態分佈於細胞質中,少數后含物還可以細胞壁增厚的狀態存在,如馬錢子、咖啡和柿子等種子的胚乳細胞,其增厚的細胞壁就是由半纖維素形成的。后含物的貯藏部位和組分隨植物生理類型的不同而各異,甜菜的肉質根及蔥的球莖鱗片細胞液內溶有醯胺、蛋白質和糖;馬鈴薯塊莖和許多其他根狀莖的薄壁組織細胞液內溶有醯胺和蛋白質,而在細胞質內則含有澱粉;其他如菜豆、豌豆子葉的薄壁組織細胞或細胞質內則存有蛋白質和澱粉等。貯藏薄壁組織中的物質積累狀況隨植物生理活動的變化而改變,木本植物的莖和根中,澱粉的沉積隨季節而變化,但在塊莖、球莖和根狀莖等貯藏器官中,當貯藏物質轉移到生長的器官以後,它們的原生質體就死亡。
貯藏的后含物中,澱粉的分佈廣泛存在於皮層、髓、輸導組織、肉質葉(鱗莖的鱗片)、根狀莖、果實、子葉以及種子胚乳的薄壁組織細胞中。除有機物質外,貯藏薄壁組織細胞也貯藏無機的礦物質,如草酸鈣、碳酸鈣、二氧化硅等各種不同種類的結晶體。有些薄壁細胞產生晶體后原生質體仍然保存,有的則在晶體發育后即死亡。
貯藏的營養物質主要作為植物本身進一步發育或繁殖後代時的能源,這在種子、塊根以及塊莖植物的發育中,表現尤為明顯。常常是幼苗產生后,這些器官中所貯藏的物質也隨之轉化與分解。
貯水薄壁組織
貯水薄壁組織是細胞中貯藏有豐富水分的薄壁組織。細胞較大型,具有一個富含水分或粘性汁液的大液泡,因而迫使細胞質、細胞核僅呈一薄層緊貼著細胞壁。粘性汁液遇水膨脹,有增加細胞吸收與保水的能力。貯水薄壁組織細胞還可存在於原生質體中和細胞壁上。許多旱生的肉質植物,如仙人掌、蘆薈、龍舌蘭、景天、松葉菊等的光合器官中,都有這種缺乏葉綠素而充滿水分的貯水薄壁組織細胞,它們排列成行,也能象柵欄狀細胞那樣延長。具有這種貯水薄壁組織的肉質植物,可以適應沙漠、石灘等少水地區的乾旱環境。
植物的地下貯藏器官里,一般沒有單獨的貯水薄壁組織,但在含澱粉及其他營養物質的薄壁組織細胞中,含水量也可達很高。例如馬鈴薯的塊莖就貯有供其在空氣中發芽,以及塊莖開始生長時所需的水分。高度含水不僅是地下貯藏器官的特點,也是氣生莖及氣生芽肉質膨大部分的特點。
通氣薄壁組織
通氣薄壁組織是薄壁細胞間隙很發達以保證空氣流通的一類薄壁組織。葉肉中的海綿組織與水生植物(如菖蒲、燈心草等)根、莖的皮層內的通氣薄壁組織最為典型。葉肉通氣薄壁組織的細胞間隙中,其空氣所佔的體積約為葉肉體積的7.7~71.3%。水生被子植物的通氣薄壁組織尤其發達,在體內形成一個相互貫通的通氣系統。使葉營光合作用而產生的氧氣能通過通氣系統進入根中。因為細胞間隙中充滿了空氣,也可增強水生植物的浮力和支持力。通氣薄壁組織的這種結構與功能的統一,是植物長期適應、進化的結果。
通氣薄壁組織中,細胞間隙的形成方式有兩種:①裂生細胞間隙——相鄰細胞細胞壁的直接連接處彼此裂開或不同程度的分離而形成。細胞分裂面與莖或葉柄的縱軸平行,與初發生的空隙表面垂直,因此這些間隙常為許多細胞所圍繞。根、莖的皮層以及葉肉海綿組織中的薄壁細胞間隙都屬這一類。在水生植物及單子葉植物,如菖蒲、燈心草、伊樂藻等的莖、葉中,細胞間隙尤為發達。②溶生細胞間隙——主要是由於形成細胞間隙的細胞在生長過程中相繼毀壞、自溶,從而出現了大的空腔而成。它常見於皮層及髓的囊狀或管狀組織,如玉米、木賊及莎草科植物的根中。
軸向薄壁組織(axialparenchyma)
由紡錘形原始細胞分生出的,一種縱行成串的磚形薄壁細胞組成。特點是具單紋孔,形體較短。在木質部的薄壁組織,稱為木薄壁組織。木薄壁組織在針葉林中含量少或缺。約佔總體積1.5%,但在羅漢松科、杉科和柏科等木材中相比之下。較發達,為該類木材的重要識別特徵。其它針葉材含量少或無。針葉材木薄壁細胞在橫斷面上呈方形或長方形。壁薄,且胞腔內含有深色樹脂故又稱樹脂細胞。橫切面呈褐黑色,易辨別。在縱切面上常成一串,可達幾厘米長,其兩端細胞比較尖削。薄壁細胞與管胞的區別:壁薄、單紋孔、短、端壁水平(因單紋孔對常呈珠瘤狀起,對鑒別木材有一定的價值)。因薄壁細胞腔內含有樹脂、芳香油,如杉木、圓柏、柏木,可浸提出杉、柏木油,它則可使木材具有特殊的香味。而且使木材耐久性大為提高。分佈類型:①星散型:星散分佈於年輪之中,如杉木。②間位型:連接成斷續的切線狀,弦向排列,如柏木。③輪界型:分佈於年輪外側,如鐵杉、黃杉。三、木射線系組成針葉材的主要分子之一。存在於所有針葉材中。但含量僅占木材總體積7%左右。在顯微鏡下觀察,木射線為多數的細胞組成,呈輻射狀,每個單獨的細胞,稱為射線細胞。木射線大多由薄壁細胞組成。這種構成木射線的單個薄壁細胞,我們叫射線薄壁細胞。但也兼有厚壁細胞組成的這種厚壁cell叫射線管胞。它是木材唯一的橫向厚壁細胞。存在於松屬、雲杉屬、落葉松屬、雪松屬、銀杉屬、黃杉屬等屬木材中。另外在某些樹種(具橫向樹脂道)中,木射線中有泌脂細胞。木射線:本由木質部中的許多射線細胞相互連續聚合而成的組織。1、木射線的種類根據其在弦切面上的形態,可分為兩種。①單列木射線:僅有一列(或偶有2列)木射線細胞所構成的射線。如冷杉、杉木、柏木等。②紡錘形木射線:多列木射線或在木射線的中央,由於橫向樹脂道的存在而使木射線呈紡垂形。常見於具橫向樹脂的樹種,如松、雲杉、落葉松、銀杉、黃杉等屬樹種中。2、木射線的組成針葉樹材木射線主要為射線薄壁細胞所組成。但偶而亦有上述講的射線管胞夾雜在某些針葉樹材射線之內。①射線管胞:是木射線中與木紋成垂直方向的橫向管胞為松科木材的特徵。胞壁具具緣紋孔。但少而不。長約為軸向管胞的1/30。長寬比為5—10。腔內無樹脂,它一般多出現於射線,薄壁細胞的上下邊緣,成1-2例,偶而混生於射線薄壁細胞之間。射線管胞內壁,有些樹種有鋸齒狀加厚。如馬尾松,油松等。有的內壁平滑,如紅松、華山松、白皮松等。在松屬中具齒狀加厚的屬硬松類,不具齒狀加厚的屬軟松類。齒狀加厚的有無、程度為鑒別木材的特徵之一。射線管胞的內壁鋸齒狀加厚,以晚材最為發達。徑切面上最易觀察。另射線管胞內壁也可能在有螺紋加厚。如某些雲杉及黃杉。②射線薄壁細胞:是組成木射線的主體,為橫向生長的薄壁組織,為矩形;長方形或略不規則。射線薄壁細胞與射線管胞之間的紋孔對為半具緣紋孔對。A、水平壁與端壁:射線薄壁細胞水平壁的厚薄為識別木材的依據之一。(如射線薄壁細胞的水平壁比鄰近的管胞壁薄。就認為該木射線的細胞壁薄)。水平壁薄是南洋杉、羅漢松屬及柏科少數屬的特徵。比起水平壁厚薄;水平壁上有無紋孔,則較重要(對木材識別而言)。在松科中一些木材的射線薄壁細胞的水平壁上,具顯著的紋孔,在晚材部分最易觀察。如雲杉、落葉松、黃杉、鐵杉、冷杉、雪松、油杉及金錢松等。但杉科、南洋杉科、紅豆杉科、三尖杉科、羅漢松科、及柏科的水平壁上則無顯著的紋孔。如圖3-13,P76,水平壁全無紋孔的為羅漢松及南洋杉兩科。端壁的節狀加厚(珠瘤狀起)也是鑒別木材的特徵之一。松科其中的軟松系樹種及刺柏屬。翠柏屬等部分樹種,均具節狀加厚。另射線薄壁細胞的水平壁和端壁的交接處。具有凹痕,如紅豆杉、圖3-4P27。它是指徑切面上射線薄壁細胞四隔的凹痕,除南洋杉科,余大多有。下面我們談談木射線中一個很重要的部分。B、交叉場紋孔的類型:交叉場:在徑切面上木射線薄壁細胞與軸向管胞相交的平面(grossfieldpitting),一般指早材部分。在此平面區域內的紋孔,稱為交叉場紋孔。它是針葉樹材最重要的識別特徵之一。其區內紋孔的形狀、數目對木材識別和分類均具重要意義。現一般將其類型分為五類:①窗格狀:單紋孔,形狀大呈窗格狀。每交叉場內有1-3個。為很多松屬樹種的特徵。如馬尾松、樟子、松、雲南松等。亦存在於羅漢松科和杉科的某些屬木材中。但不甚典型。②雲杉型:具窄而稍外延或內涵的紋孔口,形狀小。為雲杉、落葉松、黃杉和粗榧屬木材典型而明顯的特徵。在南洋杉科、羅漢松科、杉科的杉屬及雪松等木材中;有時與其主要類型紋孔同時出現。③柏型:紋孔口內含,較雲杉型稍寬。其長軸隨位置而變。從垂直位置到水平。紋孔數一般1-4個。為柏科的特徵。但雪松、鐵杉屬及油杉屬的木材中也可發現。同南洋杉科通常數目較多,排列不規則。④杉型:為卵圓至圓形的內含紋孔。紋孔口較柏型大寬。長軸與紋孔緣一致。為杉科大部分木材所具有的特徵。但在冷杉屬、羅漢松科及雪松屬木材中亦常發現。不過,常與其主類型紋孔聚在一起。⑤松型:較窗格狀小,單紋孔或具狹緣。無一定形狀。具狹緣時,與杉型相似,但其紋孔口的兩端較尖,紋孔的大小不一。常見於松屬木材。如白皮松。弦切面上,射線細胞多呈圓形,卵圓形或近圓形,射線兩端比中部的長。在高木射線中比在低木射線中的長。木射線的高度:以弦切面上的細胞個數或將整個高度以mm計量。針葉材木射線一般不高。平均為10—15個細胞高,在我國最高的為水杉,達60個,而較低的如柳杉,只有幾個。
吸收薄壁組織
吸收薄壁組織是具有吸收和傳導植物體內水分、無機鹽及有機養料功能的薄壁組織。根尖的表皮是吸收水分和無機鹽的吸收薄壁組織,尤其是根毛區的許多表皮細胞的外壁向外凸起形成根毛,更有利於物質的吸收。禾本科植物胚的盾片與胚乳相接處的上皮細胞,是吸收有機養料的吸收組織,在種子萌發時,可吸取胚乳的營養供胚胎生長發育之需。
除了上述七種類型外,20世紀60年代,藉助於電子顯微鏡技術,還發現了一種特化的薄壁組織細胞,稱為傳遞細胞。其細胞壁向胞腔內突入,形成許多指狀或鹿角狀的不規則突起,使質膜的表面積增加,並且富有胞間連絲,有利於物質的運送傳遞。這類細胞多分佈在植物體內溶質大量集中、短距離運輸頻繁的部位,如葉脈末端輸導組織的周圍,成為葉肉和輸導組織之間物質運輸的橋樑。在生殖器官中,傳遞細胞常有許多不同的形態。
其他如花上的蜜腺、松柏類植物樹脂道周圍的上皮細胞等常有濃厚的原生質體、豐富的核糖體及高爾基器等,雖然它們也屬薄壁組織,由於它們產生特殊的分泌物,故列為另一類結構——分泌結構。