共找到2條詞條名為雙受精的結果 展開
- И·Н·戈羅然金首先發現的現象
- 書籍
雙受精
И·Н·戈羅然金首先發現的現象
雙受精(double fertilization)是指被子植物中的單子葉植物的雄配子體形成的兩個精子,一個與卵融合形成二倍體的合子,另一個與中央細胞的極核(通常兩個)融合形成初生胚乳核的現象。雙受精後由合子發育成胚,中央細胞發育成胚乳。
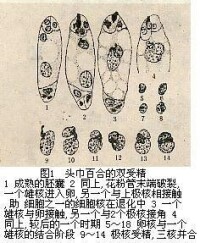
頭巾百合的雙受精
被子植物胚囊中極核同卵一樣受精產生了具有父本和母本遺傳性的通常是三倍體的胚乳,由這種胚乳“哺育”胚可能使後代更加鞏固它雙親的特性並更富有生命力,因此有人認為雙受精是被子植物繁盛的一個重要原因。
在這一段時期,發生的事態有:
⑴花粉萌發和花粉管在花柱中生長
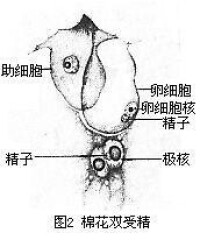
棉花雙受精
⑵花粉管進入胚囊和釋放內容物
花粉管伸入子房后,沿子房內壁或胎座繼續生長,直達胚珠,經珠孔進入胚珠,最後到達胚囊。關於花粉管沿一定道路生長的現象,一般認為是受化學物質的吸引,多數學者認為這種化學物質是由助細胞分泌的。花粉管進入胚囊后的途徑也有各種說法,如穿入一個助細胞、從卵與一助細胞之間或卵與胚囊壁之間通過等等。經過對十多種植物,如棉花、玉米、矮牽牛等的電子顯微鏡研究,證實花粉管是從一個助細胞進入的,而且進入的那個助細胞常常是在花粉管進入之前解體。花粉管就在此助細胞的位置釋放出內容物。通過電子顯微鏡觀察發現有些植物的花粉管的末端(如矮牽牛、薺菜)或末端一側(如棉花)具有一孔,因此推測花粉管中的精子和其他內容物是通過此孔釋放的。
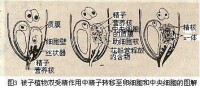
被子植物雙受精圖示
⑴雄配子與雌配子是如何併合為一體的
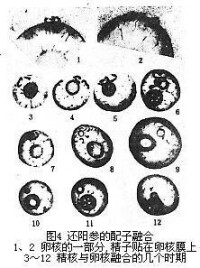
還陽參的配子融合
⑵精子的細胞質是否參與融合
按W·A·詹森的精子入卵模式,只是精子核轉移至雌性細胞中。實際上在被子植物的受精作用中,精子的細胞質是否參加迄今尚無定論。應用電子顯微鏡的研究,也得出兩種不同的結論:如在棉花的受精卵內看不到精子的細胞質,推測只有核參加受精,但在紅萼月見草的合子中則發現有精子的質體;在白花丹受精后的卵和中央細胞中也發現有精子的線粒體和質體,表明細胞質也參與融合。
⑶核的融合
配子融合的最後一步。1933年,前蘇聯植物胚胎學家E·H·格拉西莫娃·納瓦希娜和C·Γ·納瓦申對一種菊科植物——還陽參受精作用中精核與卵核的融合作了相當詳細的描述並繪有精美的圖(圖4)。根據1979年中國植物胚胎學家胡適宜等對向日葵、小麥、棉花和紫萼幾種植物所作的詳細觀察,證明被子植物雄核與雌核融合的基本程序是:①雄性核與雌性核接觸;②雄性核和雌性核之間的核膜融合;③精子核的染色質在卵核內分散和出現雄性核仁;④雄性核的染色質與雌性核的染色質混合;⑤雄性和雌性的核仁融合為一個大的核仁(圖5、6)。精核同極核(或次生核)的融合過程與精核同卵核的融合程序基本相同。
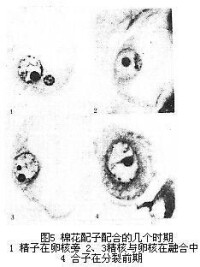
棉花配子配合的幾個時期
傳粉至受精間隔期的長短還受環境因素,特別是溫度的影響。溫度對花粉萌發和花粉管的生長速度有直接的影響。每種植物花粉萌發和花粉管生長都有自己最適的溫度,低溫可使萌發和生長受抑制,從而延長花粉管到達胚囊的時間。
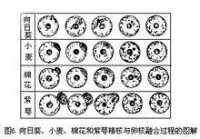
向日葵等精核與卵核融合過程圖解
在種子植物受精作用中存在不同的核融合方式。詳細研究過的幾種松科植物,如油松、白皮松和歐洲黑松等,表現為精子核進入卵核后,各自獨立形成雄性及雌性的染色體組和自己的紡錘體,真正的融合是在受精卵有絲分裂之後;而詳細研究過的幾種被子植物,向日葵、小麥、棉花和紫萼,是雄性核和雌性核的染色質混合后才分裂。

配子融合的類型
在受精前和受精后卵細胞核中脫氧核糖核酸(DNA)的含量,根據孚爾根反應方法的測定是有變化的。應用這種方法已經確定,卵細胞在發育早期是強的孚爾根正反應,而達到成熟時變為弱的正反應或負反應。受精以後形成的合子中是正的孚爾根反應。對卵細胞在發育過程中所表現的孚爾根反應強弱的變化是否代表了核中DNA含量的變化曾有爭論。20世紀70年代以來,經熒光顯微鏡和紫外光顯微分光光度計研究的結果,證明核不顯示孚爾根反應不一定缺少DNA或DNA分子的核苷酸,也就是說卵細胞核在任何時期都含有DNA。
在正常的情況下,一個胚囊只接受兩個精子,但是有時由於不止一個花粉管進入或是由於一個花粉管中形成不止一對精子,於是胚囊中出現有多於一對精子的現象。胚囊中額外精子的存在將有可能引起下面兩種異常受精作用:①多精入卵。有兩種不同的情況:一種是進入卵中多餘的精子在卵中退化,另一種是附加的精子也與卵核融合,這種異常的受精形式只在極少數植物中有細胞學的證據。例如在喜馬鳳仙花中根據在受精的卵核內看到兩個精子核的輪廓,認為是雙精子受精的。②額外的精子與胚囊中的其他細胞受精。這種現象曾在一種慈菇屬植物(Sagittariagraminea)報道過。這種植物的助細胞形態上與卵細胞相似,有兩個花粉管進入胚囊並放出額外的兩個精子,在受精后的胚囊中有時看到3個原胚存在,並有兩個花粉管在胚囊的上部。因此推測助細胞與進入的額外精子受精而產生了多胚。在還陽參中曾觀察到有些胚囊中含有2~5個卵細胞和有時一個花粉管不止兩個精子進入胚囊的情況,以及在受精后的胚囊中存在兩個受精卵在分裂的現象,推測有可能發生多於一個卵受精。

基本信息
- 中文名
- 雙受精
- 外文名
- double fertilization