神經元細胞
神經元細胞
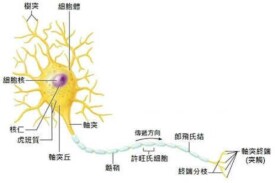
長突起的細胞,它由細胞體和細胞突起構成。細胞體位於腦、脊髓和神經節中,細胞突起可延伸至全身各器官和組織中。細胞體是細胞含核的部分,其形狀大小有很大差別,直徑約4~120微米。核大而圓,位於細胞中央,染色質少,核仁明顯。細胞質內有斑塊狀的核外染色質(舊稱尼爾小體),還有許多神經元纖維。細胞突起是由細胞體延伸出來的細長部分,又可分為樹突和軸突。每個神經元可以有一或多個樹突,可以接受刺激並將興奮傳入細胞體。每個神經元只有一個軸突,可以把興奮從胞體傳送到另一個神經元或其他組織,如肌肉或腺體。
人的大腦不同於身體的任何器官,因為出生后,它還沒有發育完全。它會在出生后的最初幾個月或幾年中,隨著他的經歷和外界刺激,繼續發育。出生時人會有數以千億計的腦細胞,當他們鏈接起來(也就是形成樹突),才可以傳導信息。如果這些神經元沒有被鏈接,是會漸漸消失的。
所以,大腦的生長,大部分原因是樹突數量和密度增加的結果。樹突在大腦中是負責接受信息的角色,從出生到4歲前,樹突的密集程度明顯加大,樹突越多說明接受信息的能力越強,人也就越聰明。
根據細胞體發出突起的多少,從形態上可以把神經元分為3類: (1)假單極神經元,胞體近似圓形,發出一個突起,在離胞體不遠處分成兩支,一支樹突分佈到皮膚、肌肉或內臟,另一支軸突進入脊髓或腦。 (2)雙極神經元,胞體近似梭形,有一個樹突和一個軸突,分佈在視網膜和前庭神經節。 (3)多極神經元,胞體呈多邊形,有一個軸突和許多樹突,分佈最廣,腦和脊髓灰質的神經元一般是這類。根據神經元的機能,可分為感覺(傳入)神經元,運動(傳出)神經元和聯絡(中間)神經元3種。神經元的功能總體來說是:受到刺激后能產生興奮,並且傳導興奮。
神經元的胞體(soma)在於腦和脊髓的灰質及神經節內,其形態各異,常見的形態為星形、錐體形、梨形和圓球形狀等。胞體大小不一,直徑在5~150μm之間。胞體是神經元的代謝和營養中心。
胞體的結構與一般細胞相似,有核仁、細胞膜、細胞質和細胞核。
(l)細胞膜:胞體的胞膜和突起表面的膜,是連續完整的細胞膜。除突觸部位的胞膜有特優的結構外,大部分胞膜為單位膜結構。神經細胞膜的特點是一個敏感而易興奮的膜。在膜上有各種受體(receptor)和離子通道(ionic chanel),二者各由不同的膜蛋白所構成。形成突觸部分的細胞膜增厚。膜上受體可與相應的化學物質神經遞質結合。當受體與乙醯膽鹼遞質或γ-氨基丁酸遞質結合時,膜的離子通透性及膜內外電位差發生改變,胞膜產生相應的生理活動:興奮或抑制。
(3)細胞質:位於核的周圍,又稱核周體(perikaryon)其中含有發達的高爾基複合體、滑面內質網,豐富的線粒體、尼氏體及神經原纖維,還含有溶酶體、脂褐素等結構。具有分泌功能的神經元,胞質內還含有分泌顆粒,如位於下丘腦的一些神經元。
l)尼氏體(Nissl body):又稱嗜染質(chromophil substance),是胞質內的一種嗜鹼性物質,在一般染色中島被鹼性染料所染色,多呈斑塊狀或顆粒狀。它分佈在核周體和樹突內,而軸突起始段的軸丘和軸突內均無。依神經元的類型和不同生理狀態,尼氏體的數量、形狀和分佈也有所差別。典型的如脊髓前角運動神經元,尼氏體數量最多,呈斑塊狀,分散於神經原纖維之間,有如虎皮樣花斑,故又稱虎斑小體(tigroid body)。而在脊神經節神經元的胞質內,尼氏體呈顆粒狀,散在分佈。電鏡下,尼氏體是由許多發達的平行排列前粗面內質網及其間的遊離核糖體組成。神經活動所儒的大量蛋白質主要在尼氏體合成,再流向核內、線粒體和高爾基複合體。當神經元損傷或中毒時,均能引起尼氏體減少,乃至消失。若損傷恢復除去有害因素后,尼氏體又可恢復。因此,尼氏體的形態結構可作為判定神經元功能狀態的一種標誌。
2)神經原纖維(neurofibril):在神經細胞質內,存在著直徑約為2~3μm的絲狀纖維結構,在銀染的切片體本可清晰地顯示出呈棕黑色的絲狀結構,此即為神經原纖維,在核周體內交織成網,並向樹突和軸突延伸,可達到突起的未消部位。在電鏡下觀察,神經原纖維是由神經絲甜神經微管集聚成束所構成。神經絲(neurofilament)或稱神經細絲,是直徑約為10nm細長的管狀結構,是中間絲的一種,但與 其他細胞內的中間絲有所不同。在電鏡高倍放大觀察。可見神經細絲是極微細的管狀結構,中間透明為管腔,管壁厚為3nm,其長度特長,多集聚成束。分散在胞質內,也延伸到神經元的突起中。神經絲的生理功能是參與神經元內的代謝產物和離子運輸流動的通路。神經微管(neurotubule)是直徑約25nm的細而長的圓形細管,管壁厚為5nm,可延伸到神經元的突起中,在胞質內與神經絲配列成束,交織成網。其生理功能主要參與胞質內的物質轉運活動,接近微管表面的各種物質流速最大,微管的表面有動力蛋白(dynein),它本身具有ATP酶的作用,在ATP存在狀態下,可使微管滑動,從而使微管具有運輸功能。此外,還有較短而分散的微絲。微絲(microfilament)是最細的絲狀結構,直徑約5nm,長短不等,集聚成束,交織成網,廣泛的分佈在神經元的胞質和突起內,其主要功能具有收縮作用,適應神經元生理活動的形態改變。神經絲、微管、微絲,這三種纖維,構成神經元的細胞骨架(cytoskeleton),參與物質運輸,在光鏡下所顯示僅是神經絲和神經微管形成的神經原纖維。其生理功能主要參與胞質內的物質轉運活動,接近微管表面的各種物質流速最大,微管的表面有動力蛋白(dynein),它本身具有ATP酶的作用,在ATP存在狀態下,可使微管滑動,從而使微管具有運輸功能。此外,還有較短而分散的微絲。微絲(microfilament)是最細的絲狀結構,直徑約5nm,長短不等,集聚成束,交織成網,廣泛的分佈在神經元的胞質和突起內,其主要功能具有收縮作用,適應神經元生理活動的形態改變。神經絲、微管、微絲,這三種纖維,構成神經元的細胞骨架(cytoskeleton),參與物質運輸,在光鏡下所顯示僅是神經絲和神經微管形成的神經原纖維。 3)脂褐素(lipofuscin):常位於大型神經無核周體的一側,呈棕黃色顆粒狀,隨年齡增長而增多,經電鏡和組織化學證實為次級溶酶體形成的殘餘體(residual body),其內容物為溶酶體消化時殘留的物質,多為異物、脂滴或退變的細胞器。某些神經元,如下丘腦,具有內分泌功能的分泌神經元(secretoryneuron),腦體內含直徑IO0~30Onm的分泌顆粒,顆粒內含肽類激素(如加壓素、催產素等)。
神經元的突起是神經元胞體的延伸部分,由於形態結構和功能的不同,可分為樹突和軸突。
樹突(dendrite)
是從胞體發出的一至多個突起,呈放射狀。胞體起始部分較粗,經反覆分支而變細,形如樹枝狀。樹突的結構與腦體相似,胞質內含有尼氏體,線粒體和平行排列的神經原纖維等,但無高爾基複合體。在特殊銀染標本上,樹突表面可見許多棘狀突起,長約0.5~1.0μm,粗約0.5~2.0μm,稱樹突棘(dendritic spine),是形成突觸的部位。一般電鏡下,樹突棘內含有數個扁平的囊泡稱棘器(spine apparatus)。樹突的分支和樹突棘可擴大神經元接受刺激的表面積。樹突具有接受刺激並將衝動傳入細胞體的功能。
軸突(axon)
每個神經元只有一根胞體發出軸突的細胞 質部位多呈賀錐形,稱軸丘(axon hillock),其中沒有尼氏體,主要有神經原纖維分佈。軸突自胞體伸出后,開始的一段,稱為起始段(initial segment),長約 15~25μm,通常較樹突細,粗細均一,表面光滑,分支較少,無髓鞘包卷。離開胞體一定距離后,有髓鞘包卷,即為有髓神經纖維。軸突末端多呈纖細分支稱軸突終未(axon terminal),與其他神經元或效應細胞接觸。軸突表面的細胞膜,稱軸膜(axolemma),軸突內的胞質稱 軸質(axoplasm)或軸漿。軸質內有許多與軸突長袖平行的神經原纖維和細長的線粒體,但無尼氏體和高爾基複合體,因此,軸突內不能合成蛋白質。軸突成分代謝更新以及突觸小泡內神經遞質,均在胞體內合成,通過軸突內微管、神經絲流向軸突末端。神經元樹突的末端可以接受其他神經傳來的信號,並把信號傳給神經元,因此是傳入神經的末梢。而軸突的分枝可以把神經傳給其他神經元或效應器,因此是傳出神經的末梢。電鏡下,從軸丘到軸突全長可見有許多縱向平行排列的神經絲和神經微管,以及連續縱行的長管狀的滑面內質網和一些多泡體等。在高倍電鏡下,還可見在神經絲、神經微管之間均有極微細纖維網路連接,這種橫向連接的極細纖維稱為微小梁(microtrabecula)起支持作用。軸突末端還有突觸小泡。軸突運輸(axonal transport)神經元的胞體和軸突在結構和功能上是一個整體,神經元代謝活動的物質多在胞體形成,神經元的整體生理活動物質代謝是由軸漿不斷流動所實現。研究證明:神經元胞質自胞體向軸突遠端流動,同時從軸突遠端也向胞體流動。這種方向不同、快慢不一的軸質雙向流動稱為軸突運輸。從胞體向軸突遠端的運輸,由於運輸方向與軸質流動的方向一致故稱為傾向運輸(antrograde transport),這種運輸有快慢之分:快速運輸,其速度為每天200~500mm,是將神經元胞體合成的神經遞質的各類小泡和有關的酶類等經長管狀的滑面內質網和沿微管表面流向軸突末端,待神經衝動時釋放。慢速運輸也稱軸質流動(axoplasmic flow),其速度為每天1~4mm,主要是將神經元胞體合成的蛋白質,不斷地向軸突末端流動,以更新軸質的基質、神經絲以及微管等結構蛋白質。逆向運輸(retrograde transport)是軸突末端代謝產物和軸突末端通過人胞作用攝取的蛋白質、神經營養因子以及一些小分子物質等由軸突末端運向胞體,運輸方向與軸質流動相反,故稱為逆向運輸,速度為每天l~4mm,這種運輸主要是由多泡體實現。多泡體是一個大泡內含許多小泡,小泡內分別含有代謝產物或攝入的神經營養因子。代謝產物被逆向運輸至胞體后,經溶酶體的作用,可分解消化更新,神經營養因子到胞體后,可促進神經元的代謝和調節神經元的生理功能。不論是順向或逆向運輸,均由線粒體提供ATP供能所實現。在某種原因而感染時,有些病毒或毒素由逆向運輸,轉動到神經元的腦體內而致病。軸突運輸是神經元內各種細胞器生理功能的重要體現。軸突的主要功能是將神經衝動由胞體傳至其他神經元或效應細胞。軸突傳導神經衝動的起始部位,是在軸突的起始段,沿軸膜進行傳導。
基本信息
- 中文名
- 神經元細胞
- 外文名
- Neurons
- 別名
- 神經組織
- 結構組成
- 神經系統結構和功能