血型遺傳
血型遺傳
廣義的血型泛指高等動物和人類血液中的紅細胞、白細胞、血小板以及各種血漿蛋白質的抗原型別,狹義的血型僅指紅細胞抗原的型別,後者是常用的血型定義。1924年德國學者 F.伯恩斯坦證明ABO血型分別為三個復等位基因所控制,開創了血型遺傳的研究。血型遺傳是臨床輸血和器官移植配型的理論基礎。血型作為一種遺傳性狀很少受環境的影響,因此是極好的遺傳標誌,可用於親子鑒定、疾病關聯分析和人種演化研究。
自從1900年奧地利學者Landsteiner氏發現血型以來,人們對“血型”一詞的概念,認為是人體的一種遺傳性狀。隨著醫學科學的發展,這一概念已不能完善地表明血型的含義了。因為現今的血型抗原,不僅局限於紅細胞的血型抗原系統,而且廣泛涉及到人體的各個部分,即人體各種細胞(血細胞、組織細胞)和各種體液成分的抗原抗體系統所表現出來的遺傳多態性。
ABO血型系統
血型遺傳
Lewis血型系統
1946年法國學者 A.E.穆朗特發現抗Le2抗體,1948年P.H.安德雷森發現與此對應的抗Leb抗體。Lewis血型有Le(a+b-)、Le(a-b+) 和Le(a-b-)三種表型(+表示有這抗原,-表示沒有這抗原)。Lewis抗原原來是體液里的抗原,出現在紅細胞上是附著上去的結果。
Rh血型系統
1940年K.蘭德施泰納和A.S.威納用獼猴(Macacus rhesus)的紅細胞免疫兔或豚鼠,發現所得血清可凝集約85%白人的紅細胞,這樣的人稱為Rh陽性,紅細胞不被凝集的人為Rh陰性。Rh血型涉及五種抗原,可分別用五種人體免疫血清檢出。
關於Rh血型系統的遺傳有兩種學說,目前還不能證明或排除其中任何一種。英國統計學家和遺傳學家R.A.費希爾和英國學者R.R.雷斯認為Rh血型為三個緊密連鎖的座位所控制三個座位構成一個基因複合體,每個座位上有一對等位基因,稱為C和c、D和d及E和e,一共可以構成八種Rh基因複合體CDE、CDe、CdE、Cde、cDE、cDe、cdE和cde。這八種Rh基因複合體構成36種基因型和18種表型。五種抗血清可以用來檢出除 d以外的五種相應抗原,但是至今沒有發現抗d血清,因此d基因是假設的。
血型遺傳
Xg血型系統
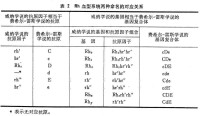
1962年J.D.曼等用一個多次輸血的病人的血清查出一種新的紅細胞抗原Xg2。這一抗原受X染色體短臂上Xg座位控制,有兩個等位基因Xg2和Xg,Xg是無效等位基因,表型可以是Xg(a+)或Xg(a-)。Xg2是迄今所知唯一X連鎖的紅細胞抗原。如果父親是Xg(a+),母親是Xg(a-),則所有女兒都將是Xg(a+),所有兒子都將是Xg(a-)。如果父親是Xg(a-),母親是Xg(a+),則子女可能都是Xg(a+),也可能半數是Xg(a+),半數是Xg(a-)。Xg血型系統的基因型和表型見表3。
其他血型系統
迄今已經發現的血型系統有十幾個,例如MNSs、Kell、p等。
一般來說血型是終生不變的。人類的血型通常分為 A 、 B 、 O 和 AB 四種。血型遺傳藉助於細胞中的染色體。人類細胞中共有 23 對染色體,每對染色體分別由兩條單染色體組成,其中一條來自父親,另一條來自母親。染色體的主要成份是決定遺傳性狀和功能的脫氧核糖核酸,即人們常說的DNA 。 DNA 可分為很多小段,每一小段都具有專一的遺傳性狀及功能,這些小段稱為基因。一對染色體中兩條單染色體上相同位置的 DNA 小片段,稱為等位基因。
ABO 血型系統的基因位點在第 9 對染色體上。人的 ABO 血型受控於 A 、 B 、 O 三個基因,但每個人體細胞內的第 9 對染色體上只有兩個 ABO 系統基因,即為 AO 、 AA 、 BO 、 BB 、 AB 、 OO 中的一對等位基因,其中 A 和 B 基因為顯性基因, O 基因為隱性基因。
紅細胞血型抗原是一類糖蛋白;抗原特異性由糖蛋白的糖基結構決定。血型基因產物是一些專一性的糖基轉移酶,它們分別催化血型抗原前體特定部位的糖基化反應,使形成相應的特異性抗原。ABO血型中的A抗原和B抗原的前體是H物質,H物質的形成受H基因控制。基因型HH和Hh的個體中有H基因產物 L-岩藻糖轉移酶-1。這種酶催化在糖蛋白前體物質末端的半乳糖上接上一個L-岩藻糖,使它轉變為H物質。ABO座位上IA基因的產物是N-乙醯-D-半乳糖胺轉移酶,IB基因的產物是D-半乳糖轉移酶,前者催化在H物質上加上N-乙醯-D-半乳糖胺使它成為A抗原,後者催化在H物質上加上D-半乳糖使它成為B抗原。O型的人沒有IA基因和IB基因,因此他們的紅細胞只有H物質,不能被抗A和抗B血清凝集,但是能被植物血凝素凝集,因此ABO血型有時也稱為ABH血型。屬於隱性純合體hh的個體中沒有L-岩藻糖轉移酶-1,不能產生H物質,紅細胞不能被植物血凝素凝集,這是一種特殊的O型,稱為孟買型。孟買型的人如有IA基因或IB基因,那麼可以有IA基因和/或IB基因編碼的相應的糖基轉移酶,但是沒有A抗原和B抗原。
Lewis抗原與A抗原和B抗原有密切關係。Lewis座位上的Le基因的產物是L-岩藻糖轉移酶-2,它催化在糖蛋白前體物質上加上一個岩藻糖,使它成為Le2抗原;或者催化在H物質上加上一個岩藻糖,使它成為Le2抗原。
ABO血型又有分泌型和非分泌型之分。在分泌型者的唾液等分泌物中可檢出相應的A、B、H等物質。在非分泌型者中不能檢出這類物質。這一性狀為Se基因所控制,Se Se和Se se是分泌型,se se是非分泌型。
猿類紅細胞有ABO抗原,血清中有相應的抗體,唾液中有A、B、H物質。狒狒和大多數猴類的紅細胞沒有ABO抗原,但唾液中有A、B、H物質。猿、狒狒和猴的唾液中有 Lewis物質。這些事實說明在進化過程中唾液中的ABO抗原出現在先,紅細胞上的ABO抗原出現在後。
猿類紅細胞有MN血型。豚鼠抗恆河猴紅細胞血清可用來檢出人類的一種與Rh抗原有關的LW抗原。猿類、狒狒和多種猴類的紅細胞有LW抗原。猴類沒有Rh抗原,但所有猿類都有某種形態的D抗原和c抗原,E/e系列抗原只見於人類。這又說明在進化上LW基因比Rh基因古老;各種Rh基因也是在進化過程中逐漸出現的。
輸血和移植配型
在臨床輸血中如果受血者血液含有針對供血者紅細胞抗原的抗體,常導致嚴重的溶血性輸血反應。輸入的血液如含有針對受血者紅細胞抗原的抗體則並無妨礙。這是因為輸入的抗體立即被受血者的血液稀釋,並為組織細胞所吸收的緣故。因此輸血配型的原則是輸入的紅細胞不能具有受血者所缺的抗原。
臨床器官移植中選擇供者和受者時,必須使紅細胞血型相配合,其原理與輸血配型完全相同。ABO血型必須配合,Rh血型和P血型對移植物存活的影響尚無定論。在受者使用免疫抑製劑的情況下,未見其他血型系統有影響。
這是胎兒與母親紅細胞血型不配合的結果。各種血型不配合都可引起新生兒溶血症,其中以Rh血型不配合引起的溶血症最為嚴重。在中國多見的是癥狀較輕的ABO新生兒溶血症。
親子鑒定
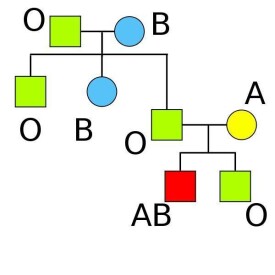
根據血型遺傳規律可排除親子關係。例如O型母親有一個A型的孩子,則B型和O型的男子不可能是這孩子的生物學父親,余類推。
某些血型基因座位在染色體上的位置已經確定,如Duffy、Rh、Scianna、Dombrock等座位在第一號染色體上,P座位在第6號染色體上,Colton和Kidd座位在第7號染色體上,ABO座位在第9號染色體上,Xg座位在X染色體上。這些血型可以作為體細胞雜交基因定位研究中的重要的遺傳標誌。
疾病遺傳研究
利用 Xg血型可確定X染色體的來源。例如特納氏綜合征女孩的性染色體是X0,假定該女孩是Xg(a+),父親也是Xg(a+),而母親是Xg(a-),那麼她的X 染色體必定來自父親。又例如克氏綜合征男孩的性染色體是XXY,假定該男孩是Xg(a+),母親是Xg(a-),那麼他必定除了Y染色體外還有一條X染色體來自父親。血型也可以作為遺傳標誌被應用於產前診斷。例如已知ABH的分泌基因 (Se)和杜氏肌營養不良症的致病基因是X染色體上兩個緊密連鎖的基因,由於重組率很低,所以只要在有該病患兒娩出史的孕婦的羊水中檢出ABH,就可判斷其胎兒可能是患兒。
群體遺傳學和人類學的研究
血型這一性狀不受環境的影響而改變,身體中即使個別體細胞的血型基因發生突變,也不致影響到個體血型的改變,也不會發生血型的選擇作用。因此血型在人群中的分佈差別、同一種血型在不同民族和不同人種中的差別以及同一民族或同一人種中各種血型的頻率都是群體遺傳學和人類學的重要研究內容。
染色體
位於細胞核中,略似棒狀,著色很深的一種小體,只有在細胞分裂時才能識別。它是由許多呈線狀排列的基因構成。
基因
是位於染色體上的一段DNA,是遺傳的基本單位,能夠表達出特定的功能,決定特定的性狀以及這些性狀的變異。
顯性基因與隱性基因
用來表示子代性狀的。當兩個相對親代結合在一起時,能夠顯示出特徵來的稱為顯性基因,反之,稱為隱性基因。例如,開黃花的植物和開白花的植物雜交后,如果子代開的花仍是黃色的,那麼,開黃花的子代是顯性,而開白花的子代是隱性。
純合子與雜合子
在一對染色體的相應位點上,具有相同的等位基因,稱為純合子;反之,稱為雜合子。
表現型與遺傳型
用抗血清能夠檢測出來的后型稱為表現型:反之,稱為遺傳型。
等位基因與復等位基因
兩個或兩個以上基因中的一個,它決定著遺傳中兩個相對的性狀,位於同源染色體的相同位點。稱為等位基因:反之,在同源染色體的相同位點,那在一個基因位點上存在兩個以上的基因時,稱復等基因。
同源染色體
二倍體細胞中染色體以成對的方式存在, 一條來自父本,一條來自母本,且形態、大小相同,並在減數分裂前期相互配對的染色體。含相似的遺傳信息。
人類在第二代里,可獲得上一代的某些特徵,這種現象叫做遺傳。人的血型是按孟德爾分離與組合規律遺傳的。就現代細胞學研究看來,細胞核內染色體,每一種生物都有一定數量和一定形狀。例如人的體細胞內有46個染色體,而在性細胞內(精子和卵子)卻只有體細胞內一半數目的染色體,即23個染色體。這是由於性細胞減數分裂把成對的同源染色體分開,分配到兩個配子中去的結果。當兩性交配后形成的受精卵,其兩性的同源染色體又相互配對,故此後產生的體細胞就是含46 個(23對)染色體了。一半來自父親,一半來自母親。染色體是負責遺傳的,每一條染色體上有很多基因,基因的排列是有一定順序的,各有其特定的位置。人的 ABO血型就是由A、B、O三個基因控制的,它們在染色體上處於同一基因位點,所以ABO基因是復等位基因。換言之,在這一位點上可是O基因,也可能是A 或B基因,三者必居其一,它們只能在這一位點上,而不能佔據其他基因位點。
體細胞上的染色體如果來自父母雙方的等位基因是相同的叫純合子(例如:有的A型染色體上的等位基因A及A):如果所含的基因內容不相同則叫雜合子(例如L 有的A型人染色體上等位基因為A及O)。換句話說,每一個體,在某位點上,有兩個而且也只有兩個等位基因,每個等位基因,來自父母各一方。如果來自父母雙方的基因是相同的,此個體即稱為純合子,如不相同,則稱為雜合子。不論是純合子還是雜合子,一對染色體上基因的總和稱為遺傳式。如上述兩種人的遺傳式分別為AA和AO。但是,遺傳下來的基因不一定都表現出來,把能夠表現出來的性狀稱為表現式。如無論遺傳為AA和AO,其表現式均為A,這就是我們平常所說的 A型。可見血型實際上是指它的表現形式而言的。血型的表現式和遺傳式其所以不同,是因為有些基因無論是純合子還是雜合子,它所控制的性狀都可以表現出來,這種基因叫顯性基因,如果這種基因所控制的性狀,只有在純合子時才表現出來,而在雜合子時不能表現出來,這種基因則稱為隱性基因。在ABO血型系統中,A 和B基因是顯性基因,而O基因則是隱性基因。例如,在一對染色體中,一個染色體帶A基因,另一個帶O基因,這個人的遺傳式為AO,但表現式為A,即是A 型,而不是O型。O型人體這是兩個染色體上體上O基因。
以此推理,ABO血型系統的遺傳式和表現如表:
表1:ABO血型及其相應的遺傳式
血型或表現式 遺傳式或基因型
A AA,AO
B BB,BO
O OO
AB AB
即表現式A,B,O和AB;遺傳式AA,AO,BB,BO,OO和AB。
ABO血型鑒定,通常只用兩種抗血清即抗A及抗B血清,就可將群體分為四種血型。
根據血型的遺傳規律,和臨床工作方例起見,配偶間所生子女的血型見表2。
表2:各種ABO配偶所生子女的血型:
婚配式 子女可能有的血型 子女不可能有的血型
| 父母血型 | 子女可能有血型及比例 | 子女不可能有血型 |
| O、O | O | A、B、AB |
| O、A | O、A (1:3) | B、AB |
| O、B | O、B (1:3) | A、AB |
| O、AB | A、B (1:1) | O、AB |
| A、A | O、A (1:15) | B、AB |
| A、B | A、B、AB、O (3:3:9:1) | — |
| A、AB | A、B、AB (4:1:3) | O |
| B、B | O、B(1:15) | A、AB |
| B、AB | A、B、AB(1:4:3) | O |
| AB、AB | A、B、AB(1:1:2) | O |
基本信息
- 中文名
- 血型遺傳
- 外文名
- Inheritance
- 別名
- ABO血型系統、Lewis血型系統
- 拼音
- xuexingyichuan
- 提出者
- 奧地利醫學家K.蘭德施泰納
- 應用
- 親子鑒定、疾病關聯分析