棘龍科
棘龍科
棘龍科(Spinosauridae)是群巨大、獨特的獸腳亞目食肉恐龍。它們是群相當大型的二足掠食動物,擁有修長、類似上龍的頭顱,上有圓錐狀的牙齒,這些牙齒有非常小的鋸齒狀邊緣,或者沒有鋸齒狀邊緣。齒骨的前部牙齒往外突出,還有著強壯發達的前肢,巨大的爪子,這些成為這群食肉動物的外表特徵。棘龍科的名稱來自於模式屬棘龍的背部帆狀物,由多條神經棘構成。這些帆狀物的功能仍在爭論中,但普遍的解釋是做為調節體溫使用、用來威嚇可能的攻擊者、或是視覺辨識物。有些古生物學家更提出,這些神經棘支撐者由肌肉、脂肪構成的隆肉。
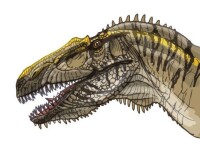
棘龍科頭部特寫
棘龍科(Spinosauridae)是群獨特的獸腳亞目恐龍。是一類獸腳類恐龍,比較有名的棘龍在中白堊紀階段是繁盛期,約14000至6500萬年前。它們是群相當大型的二足掠食動物,擁有修長、類似鱷魚的頭顱,上有圓錐狀的牙齒,這些牙齒有非常小的鋸齒狀邊緣,或者沒有鋸齒狀邊緣。齒骨的前部牙齒往外突出,成為這群動物的外表特徵。棘龍科的名稱來自於棘龍的背部帆狀物。
棘龍科的化石已在非洲、歐洲、南美洲、亞洲、澳洲、以及北美洲等地發現。其他的大型掠食性恐龍都有適合撕咬的牙齒與頜部,而棘龍科的牙齒與頜部似乎較適合捕食大型魚類。

棘龍骨架
已知最早的 棘龍科恐龍出現於侏羅紀晚期啟里莫階,約1億5500萬年前,截止2018年發現的侏羅紀晚期化石,僅限於牙齒化石。生存在東非坦尚尼亞的東非龍,是已知最早的棘龍科成員,被認為是基礎棘龍科恐龍。棘龍科繁盛於白堊紀早期的巴列姆階到白堊紀中期的森諾曼階,是非洲的頂級掠食者和次等掠食者。其中的最大型成員棘龍是到2018年為止最大的獸腳亞目食肉恐龍和體型最大的陸地食肉動物。
由於白堊紀中期森諾曼階的地質大變動,使得岡納瓦大陸解體,非洲、南美洲、澳洲的自然環境受到了嚴重破壞,大量蜥腳類和鳥臀目恐龍因為缺乏食物而死亡,作為頂級掠食者棘龍科恐龍失去了主要食物來源,也走向了衰落。各地頂級掠食者的位置漸漸地被興起的阿貝力龍科和暴龍科恐龍所取代。阿根廷的土侖階地層,曾發現過類似 棘龍科的牙齒,但2009年的研究顯示發現這些牙齒屬於鑲嵌踝類主龍。近年在中國河南省馬家村組發現一顆重爪龍亞科的牙齒,地質年代屬於桑托階中期。

棘龍科最晚的牙齒化石
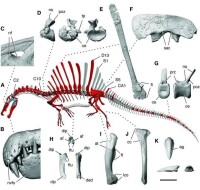
埃及棘龍最新化石
棘龍科包含兩個亞科:重爪龍亞科、棘龍亞科。棘龍亞科是由塞里諾在1998年所命名,並由湯瑪斯·荷茲(Thomas R. Holtz Jr.)等人在2004年定義為:親緣關係較接近埃及棘龍,而離沃克氏重爪龍較遠的所有物種。重爪龍亞科則是由查理格(Alan J. Charig)與Angela C. Milner在1986年所建立,它們為了新發現的重爪龍屬建立了重爪龍科與重爪龍亞科;後來重爪龍被發現與 棘龍科有關。湯瑪斯·荷茲等人在2004年將重爪龍亞科定義為:親緣關係較接近沃克氏重爪龍,而離埃及棘龍較遠的所有物種。
棘龍超科Spinosauroidea
棘龍科Spinosauridae
東非龍Ostafrikasaurus
重爪龍亞科 Baryonychinae
重爪龍 Baryonyx
脊飾龍 Cristatusaurus
魚獵龍 Ichthyovenator
似鱷龍 Suchomimus
鱷龍 Suchosaurus
棘龍亞科 Spinosaurinae
暹羅龍Siamosaurus(存在爭議)
扶綏中國上龍Sinopliosaurus fusuiensis(存在爭議)
激龍Irritator
棘龍Spinosaurus
奧沙拉龍Oxalaia
棘龍擁有獨特的帆狀物,雖然同一時期的該地區其他恐龍也可能發展出類似的背椎結構物,例如鳥腳下目的豪勇龍、蜥腳下目的雷巴齊斯龍,以及生存於南美洲的蜥腳類阿瑪加龍。棘龍的帆狀物也類似異齒龍的背帆;異齒龍是種似哺乳爬行動物,生存於恐龍時代以前的二疊紀,兩者的帆狀物並非同源演化的特徵,而是平行演化的結果。棘龍的背部結構物可能較類似駝峰,而非帆狀物。早在1915年,恩斯特·斯特莫就認為這些背部結構物支撐者脂肪構成的隆肉。在1997年,Jack Bowman Bailey最近提出棘龍、豪勇龍的背棘並非細棒,而是前後軸寬廣,不同於盤龍目的扁平背帆,而是類似巨角犀、水牛的背脊,所以棘龍的背棘應是支撐者較厚、較肥胖的結構物,而非細的帆狀物。
棘龍的帆狀物功能仍未確定;科學家們已提出數個假設,包含調節體溫、吸引異性的展示物。此外,如此明顯的背部特徵可使棘龍的外表看起來比較大,可威脅其他動物。
如果這些帆狀物擁有大量的血管,棘龍可使用帆狀物的大型表面積來吸收熱量。這顯示棘龍至少是某種程度的溫血動物,並生存於夜間氣溫低、冷,但天空通常很少雲的地區。棘龍與豪勇龍被認為為生存於早期撒哈拉沙漠的邊緣,這可解釋上述的功能。這些帆狀物也有可能用來釋放多餘的體溫,而非收集熱量。根據霍爾丹法則(Haldane's principle),與身體體積相比,大型動物反而擁有較小比例的表面積,它們釋放的熱量溫度較高,而吸收的熱量溫度較低。這些恐龍的帆狀物增加相當程度的身體皮膚面積,而且所增加體積最少。此外,如果將這些帆狀物遠離太陽方向,或是以90度迎接較冷的風,這些身處白堊紀非洲的動物將能更有效地降低體溫。之後有其他研究人員提出反對意見,認為如果棘龍有背帆,它們吸收熱量的效率,將高於散發熱量的效率。他們認為這些有長神經棘的恐龍,背部具有脂肪構成的隆肉,以儲存能量。
許多現代動物的複雜身體結構,在求偶季節時具有吸引異性功能。這些恐龍的帆狀物有相當可能性具有吸引求偶功能,類似孔雀的尾巴。斯特莫假設這些恐龍的雄性與雌性擁有不同大小的神經棘。如果屬實,這些帆狀物可能擁有耀眼的顏色,但這是完全建立於推測上的。
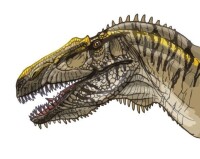
棘龍科
與其他大型掠食恐龍(例如異特龍、暴龍)相比,棘龍科的顱骨形狀、結構有非常大的不同。大部分掠食恐龍的頜部寬而高;棘龍科的頜部扁而狹窄。許多古生物學家因此推測棘龍科無法像其他獸腳類恐龍以大型、強壯的動物為食,而有不同的獵食方式。
漢斯·戴爾特·蘇伊士(Hans-Dieter Sues)等人研究激龍的頭骨,推測棘龍科是以迅速、強力的嘴部咬合,配合強壯的頸部肌肉作出快速的下咬、后縮動作,將小型獵物咬在嘴中。由於狹窄的嘴部形狀、交錯的圓錐狀牙齒,使嘴部的獵物很難脫困。
因為棘龍科具有類似現代長吻鱷的頜部與牙齒,它們過去常被視為是魚食性動物,以魚類為主食。在2007年,埃米莉·雷菲爾德(Emily Rayfield)使用重爪龍的顱骨,計算了棘龍科的生物力學結構。他們發現重爪龍的顱骨結構、咬合力,與現代長吻鱷的模式吻合;這顯示至少重爪龍亞科是以魚類為主食,而棘龍亞科似乎是多種食物來源的動物。
證據顯示棘龍科恐龍以魚類、以及許多小型到中型的動物為食,也包括了小型恐龍。曾經在重爪龍的胃部區域,發現了魚類鱗片,以及一隻年輕禽龍的被消化骨頭;另外曾在一個棘龍標本上,發現了一個魚類骨頭嵌入齒槽;此外還有一個證據顯示一隻棘龍科恐龍正吞食一隻翼龍類。有可能棘龍科是廣生性物種,以魚類、中小型動物為食。
研究人員並推測棘龍科是以魚類為主食,以避免與當地的大型獸腳類恐龍競爭相同食物來源。許多古生物學家因此推測 棘龍科有著與眾不同的獵食方式。根據分析推斷 棘龍科是以迅速、強力的嘴部咬合,配合強壯的頸部肌肉作出快速的咬合,將小型的獵物咬在嘴中,或者緊緊咬住中、大型獵物的要害部位,由於狹窄的嘴部形狀、交錯的圓錐狀牙齒配合上頜骨肌肉的力量,使得獵物很難掙脫。除此之外,與暴龍科、阿貝力龍科、鯊齒龍科不同,棘龍科的前肢很強壯發達,長著大型的指爪,被推測也是它們的主要捕食利器。
因為棘龍科具有類似現代長吻鱷的頜部與牙齒,它們過去常被錯誤地認為是純粹的魚食性動物;這顯示至少體型較小、頭部牙齒類似鱷魚重爪龍亞科是以魚類為主要食物來源,而身體結構更結實強壯、體型更大的棘龍亞科應該是多種食物來源的動物,被推測以中大型鳥臀目恐龍、小中型蜥腳類恐龍為食,但偶爾也可能食用魚類。
證據顯示棘龍科恐龍許多各種各樣的動物為食,包括了魚類、腐肉、翼;此外還有一個證據顯示一隻棘龍科恐龍正吞食一隻翼龍類。很可能棘龍科是廣生性物種,以魚類、小到大型動物為食。
研究人員並推測重爪龍亞科恐龍是次等捕食者,以魚類為主食,以避免與當地的大型獸腳類恐龍競爭相同食物來源。棘龍亞科恐龍則是其生態環境的頂級掠食者,以各種鳥臀目恐龍和小型中型蜥腳類恐龍為主食。
此外,棘龍和奧沙拉龍的頭骨很寬,而嘴前部較為狹窄,所以雙眼朝向前方,可能具備雙眼立體視覺。
