艾伯塔龍
艾伯塔龍
艾伯塔龍屬,又名亞伯達龍、阿爾伯脫龍、阿爾伯它龍、亞伯拖龍,是暴龍科艾伯塔龍亞科恐龍的一屬,生活於上白堊紀的北美洲西部,距今7000萬年前。模式種是肉食艾伯塔龍,化石是在加拿大艾伯塔省的省立恐龍公園發現,並以此省作為該屬的名字。科學家們對於艾伯塔龍有幾個種,有著不同的意見,有些科學家將平衡蛇發女怪龍列為第二個種。
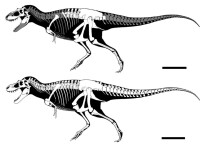
艾伯塔龍是雙足的獵食恐龍,有著大型頭部,上有很多銳利的大型牙齒,以及有兩隻手指的小型前肢。艾伯塔龍可能是位在其生態系統的食物鏈頂部。雖然在獸腳亞目中,艾伯塔龍的體型較大,平均體重2·5噸,體長6米,最大7.5米長3噸重。但仍比其著名近親暴龍更小型。
屬蜥臀目獸腳亞目的暴龍科恐龍。化石發現於北美晚白堊紀地層,因出土於加拿大的艾伯塔省而得名。

艾伯塔龍

艾伯塔龍
艾伯塔龍身長約7米,身高2米左右,體重2噸。它是一種早期霸王龍類,生活在距今約7100萬-6700萬年前,比我們熟悉的霸王龍要早三百萬年就橫行於天下。由於它體重較輕,腿部又長,因此是已知暴龍類恐龍中跑得第二快的品種,僅次於矮暴龍。艾伯塔龍的體型比一些暴龍科恐龍較小,例如特暴龍及暴龍。成年的艾伯塔龍約有9米長。有幾項利用不同方法的研究,估計成年的艾伯塔龍體重在2到3噸之間。
所有暴龍科恐龍都有相似的外貌,包括艾伯塔龍在內。艾伯塔龍是雙足動物,具有長尾巴,可平衡頭部及身軀的重量。與巨大體型相比,暴龍科的前肢是極為小型的,且只有兩指。後肢極其長,具有四個腳趾。后趾很短,只用其他三趾接觸地面,而中間的腳趾較其他為長。艾伯塔龍可能可以每小時14到21公里的時速行走。至於艾伯塔龍的幼年個體,可能可用更快速度移動。

艾伯塔龍頭骨化石
2001年的一項研究,認為艾伯塔龍牙齒的鋸齒狀邊緣,可能在進食時協助撕裂獵物的肉塊。在咬合時,牙齒鋸齒狀邊緣的形狀,可增加接觸肉塊的表面積、散布咬合時所遭受的應力。某種植龍類、盤龍目異齒龍的牙齒也帶有類似鋸齒狀邊緣,但無法散布咬合時所遭受的應力。暴龍科動物的牙齒是用來協助撕裂獵物的肉塊,而非單純切割裂物的肉塊。根據牙齒的磨損模式,顯示暴龍類在進食時會搖晃頭部,以咬下獵物。

艾伯塔龍
艾伯塔龍的模式標本是一個部份的頭顱骨,是在1884年6月被發現於艾伯塔省紅麋河邊的一個露頭,屬於馬蹄峽谷組(Horseshoe Canyon Formation)。該標本是由加拿大地質學家約瑟夫·蒂勒爾(Joseph B. Tyrrell)所帶領的加拿大地質調查局考察隊所發現的,由於缺乏識當工具,這個幾乎完整頭顱骨只有部份被保存下來。在1889年,另一個團隊也在附近發現一個較小的頭顱骨、以及一些身體骨頭。這個標本現正存放在加拿大自然博物館。在1892年,愛德華·德林克·科普將這兩個頭顱骨建立為暴風龍的其中一種(Laelaps incrassatus)。但是,暴風龍的屬名已經被一個厲蟎屬所用,所以奧塞內爾·查利斯·馬什在1877年將暴風龍改為傷龍(Dryptosaurus),科普拒絕承認馬什所更改的名稱。而在科普死後,勞倫斯·賴博在1903到1904年詳細研究這些化石時,將它們改列為傷龍的一個種(D. incrassatus)。另一古生物學家Oliver Perry Hay也使用傷龍屬的一種。稍後在1905年,奧斯本指出,D. incrassatus只是根據一些暴龍科的普通牙齒而建立的名稱,並發現這兩個頭顱骨與傷龍的模式種鷹爪傷龍有明顯差異,因此將之建立為新屬,肉食艾伯塔龍(Albertosaurus sarcophagus )。但奧斯本並沒有詳述這些化石,而是根據賴博在前一年所寫的完整敘述。這兩個發現於亞伯達省的頭顱骨,分別被列為正模標本(編號CMN 5600)、副模標本(編號CMN 5601),現正存放在渥太華的加拿大自然博物館。
艾伯塔省的紅麋河。超過一半的艾伯塔龍化石就是在此發現的。於1910年,美國古生物學家巴納姆·布郎(Barnum Brown)在紅麋河發掘出一大群艾伯塔龍的遺骸。由於大量的骨骼卻只有有限時間採集,布郎並不能採集所有標本,只是能確保從每一個個體都有採集其骨骼。七組右中骨,並兩隻在尺寸上與這些中骨不相配的獨立趾骨被採集了,且存放於紐約的美國自然歷史博物館。這顯示在石礦中最小有九隻恐龍。加拿大皇家蒂勒爾博物館於1997年再次發現該位址,並從新開展發掘的工作。這次的發掘發現十頭非常幼小的艾伯塔龍。
那原先被稱為A. arctunguis的標本亦是在近紅麋河發掘出來的,並且已存放在多倫多的皇家安大略博物館。另外六個頭顱骨及骨骼亦在艾伯塔省被發現,並存放在其他加拿大博物館。
所有肉食艾伯塔龍的化石都是於艾伯塔省的馬蹄鐵峽谷地層被發現。這個地層可以追溯至上白堊紀的麥斯特里希特階,距今約73-67百萬年前。艾伯塔龍的所有化石來自於7100-6700萬年的地層,為晚坎帕階至馬斯特里赫特階。很多其他的恐龍亦於此被發現,包括細小的獸腳亞目如似鳥龍、縴手龍及幾種馳龍科,另有不同的草食性恐龍如甲龍下目、角龍下目、厚頭龍下目及鴨嘴龍科。艾伯塔龍的化石在美國蒙大拿州、新墨西哥州及懷俄明州都有指被發現,但這並不一定是肉食艾伯塔龍,甚至根本不是艾伯塔龍。
在21世紀早期,有研究人員認為正模標本的損害狀況高,本身不能成為物種建立、鑒定比對的依據,艾伯塔龍應該被列為疑名。在2010年,湯瑪斯·卡爾(Thomas Carr)重新研究艾伯塔龍,提出正模標本、副模標本、之後發現的其他標本,都有一個相同的自衍征,上頜齶骨的后緣有個大型洞孔。根據這個研究結果,艾伯塔龍本身是個有效物種。

新墨西哥州的可能化石

艾伯塔龍
暴龍科的另一亞科為暴龍亞科,包含:特暴龍和暴龍。與較強壯的暴龍亞科相比,艾伯塔龍亞科的體型較修長,顱骨的比例較小,脛骨、蹠骨、腳部指骨較長。
艾伯塔龍的模式種是肉食艾伯塔龍(A. sarcophagus),都是由奧斯本於1905年所命名。這個學名的意思是“肉食者”,並與石棺是同一語源。已知的艾伯塔龍標本超過三十個,且有不同年齡的。
於1913年,古生物學家查爾斯·斯騰伯格(Charles Hazelius Sternberg)在艾伯塔省較早的地層發現另一個暴龍科骨骼。這個恐龍於1914年由勞倫斯·賴博(Lawrence Lambe)命名為蛇發女怪龍(Gorgosaurus libratus)。戴爾·羅素(Dale Russell)發現蛇發女怪龍與艾伯塔龍只有些微分別,遂於1970年指蛇發女怪龍是艾伯塔龍的異名。這項發現使得艾伯塔龍的時代推前了幾百萬年,並其地理位置亦向南移幾百公里。
於2003年,菲力·柯爾(Phil Currie)質疑羅素所提出的異名解釋。他審查了這兩個物種的頭顱骨,雖然這兩個屬是姊妹分類,並且有不多的分別,他仍結論出要保留兩個屬。根據柯爾的解釋,艾伯塔龍及蛇發女怪龍的相似性不及懼龍與暴龍的相似性,而懼龍及暴龍仍然分開為兩個屬。再者,在阿拉斯加州及新墨西哥州發現的幾個艾伯塔龍亞科標本也沒有被描述,所以柯爾建議將兩個屬維持分開直至有更多資料研究為止。有些學者支持柯爾的建議,但有些則不認同。
在新墨西哥州發現的艾伯塔龍亞科頭顱骨。這有可能是艾伯塔龍。

艾伯塔龍
一些其他的物種曾分類在艾伯塔龍中,但之後被確定是無效的。在1928年,威廉·帕克斯(William Parks)在紅麋河附近挖掘出一個部份骨骸(編號 ROM 807),並將之命名為艾伯塔龍的一個種(A. arctunguis),但在1970年被確認為肉食艾伯塔龍的異名。該標本存放在多倫多的皇家安大略博物館。另外,艾伯塔省還發現六個頭顱骨及骨骼,並存放在其他不同的加拿大博物館。美國的蒙大拿州、新墨西哥州、以及懷俄明州也發現了相關化石,但這些並不一定是肉食艾伯塔龍,甚至根本不是艾伯塔龍。
艾伯塔龍
在1930年,蘇聯古生物學家Anatoly Nikolaevich Riabinin將被發現於中國的一顆牙齒,命名為艾伯塔龍的一種(A. pericolosus)。這顆牙齒可能屬於特暴龍。德國古生物學家弗雷德里克·馮·休尼認為傷龍的一個種(D. incrassatus)不是疑名,並改歸類於艾伯塔龍的一個種。戴爾·羅素則認為蛇發女怪龍、艾伯塔龍是相同物種,因此將斯氏蛇發女怪龍(G. sternbergi)、蘭斯蛇發女怪龍(G. lancensis)都改為艾伯塔龍的種。斯氏蛇發女怪龍目前被認為是平衡蛇發女怪龍的幼年個體,而蘭斯蛇發女怪龍則可能是暴龍的幼年個體、或是獨立的矮暴龍屬。大纖細艾伯塔龍(A. megagracilis)的化石發現於蒙大拿州的海爾河組,是一個小型的暴龍科身體骨骼(編號LACM 28345)。在1995年,大纖細艾伯塔龍被重新命名為恐暴龍,但現在被認為只是一隻幼年暴龍。在1988年,葛瑞格利·保羅將奧氏獨龍(Alectrosaurus olseni)改歸類為艾伯塔龍的一個種,但沒有其他研究引用這個結果。在1989年,蛇發女怪龍的G. novojilovi被改歸類為艾伯塔的一種,目前被認為是特暴龍的異名。
艾伯塔龍捕獵

艾伯塔龍
大部分的艾伯塔龍化石,在死亡時為14歲,或是14歲以上。在化石中很少發現幼年個體的原因有數個,主要是因為年輕個體的骨頭較小,在化石化的過程中較不易保存下來,另一個原因則是採集化石時的偏差,挖掘人員在挖掘時,較少去發現到小型的骨頭。年輕的艾伯塔龍體型比幼年體大,但與成年體相比,仍較少被發現。有科學家提出這反映了艾伯塔龍的生命史,幼年艾伯塔龍較不容易死亡,因此化石較少被發現,而非化石化與挖掘化石的偏差。

艾伯塔龍
由巴納姆·布郎發現的艾伯塔龍骨床包含最小10頭恐龍。當中有2-3頭超過21歲,1頭較年輕的約17歲,4頭正在發育階段的在12-16歲間及1頭10歲的幼龍。另有一頭只有2歲的幼龍。
由於缺乏其他草食性恐龍遺骸及所有個體的相同保存狀況,菲力·柯爾認為這個位址並非一個像加利福尼亞州拉布雷亞瀝青坑的捕獵者陷阱。而所有動物都在同一時間死亡,估計它們是群體活動的。其他的學者對此存有懷疑,認為它們是因洪水或其他原因而被逼在一起的。
草食性恐龍,如角龍下目及鴨嘴龍科都有很多證據證實是群體活動的。然而很小發現有如此多的捕獵恐龍在同一地方。但是細小的獸腳亞目,如腔骨龍及恐爪龍都是一群被發現的,較大的異特龍及馬普龍亦都如是。有一些證據亦指其他暴龍科是群體活動,如在芝加哥菲爾德自然歷史博物館內展示的暴龍Sue在被發現時旁邊有一些碎片遺骸。在蒙大拿州的雙麥迪遜組最小有三個未命名的懼龍標本。縱然有些位址可能是臨時的或不自然的群體聚集,但這些發現都為艾伯塔龍的群體活動行為提供了證據。
柯爾亦猜測艾伯塔龍的群獵習性。細小艾伯塔龍的腳與似鳥龍的比較,估計它們是跑得最快的恐龍。年輕的艾伯塔龍可能比它們的獵物跑得更快,他猜測年輕的恐龍驅趕獵物至成年的恐龍。但是由於化石紀錄所提供的資料有限,這個假說能難去證實。
在2009年,一個研究發現數個暴龍科頭骨上的平滑洞孔,過去被當成是物種內打鬥行為的傷口,可能是由雙鞭毛生物的毛滴蟲(Trichomonas)所造成的,如同現代鳥類常被寄生生物所感染。這些恐龍科動物可能會互相咬對方,造成寄生蟲的傳染。
在2001年,布魯斯·羅斯柴爾德(Bruce Rothschild)等人發表一份獸腳類恐龍的壓力性骨折、肌腱撕裂傷研究。他們研究319個艾伯塔龍的腳掌骨骼,只發現一個骨頭帶有壓力性骨折的跡象,手部骨頭則沒有發現壓力性骨折的跡象。他們發現艾伯塔龍的壓力性骨折跡象,明顯少於肉食龍下目的異特龍。
A. arctunguis的正模標本(編號ROM 807),腸骨有個2.5厘米寬、3厘米長的深洞孔,當時研究人員並沒有將其鑒定為病徵。這標本的左腳第四蹠骨帶有骨疣。這個標本目前已改歸類於肉食艾伯塔龍。戴爾·羅素在1970年代發現的五個艾伯塔龍化石,其中兩個的肱骨帶有病徵、損傷。
干島水牛躍省立公園所發現的大量艾伯塔龍化石,大部分沒有發現病徵跡象。其中,三個腳掌趾骨被發現骨刺,導因於肌腱的不正常骨化。兩個肋骨、一個腹肋(某種真皮骨)則發現斷裂后癒合的跡象。一個成年標本的左下頜有個穿孔,以及癒合后、未癒合的咬痕。這個地區的艾伯塔龍化石,帶有病徵的機率相當低;而馬達加斯加的瑪君龍,則有19%的化石帶有病徵。
所有肉食艾伯塔龍的化石都是於艾伯塔省的馬蹄鐵峽谷地層被發現。這個地層的年代為上白堊紀的麥斯特里希特階,距今約67-73百萬年前。艾伯塔龍的所有化石來自於7100-6700萬年的地層,為晚坎帕階至馬斯特里赫特階。該地層正下方是熊掌組,是西部內陸海道的海相沉積層。在白堊紀末期,這個海道因為寒冷天氣消退,海平面因此下降,馬蹄鐵峽谷地層進而成為陸地。但是,這個海道是周期性的上升下降,曾經多次海侵馬蹄鐵峽谷地層,直到最後的海降。因此,馬蹄鐵峽谷地層有者不同的沉積層,包含離岸、岸邊、沿海等環境,例如潟湖、河口、潮坪。眾多的煤塊,似乎是古代沼澤形成的泥炭。如同該地發現的其他脊椎動物,艾伯塔龍的化石被發現於三角洲與泛濫平原沉積層內,而該地過去有過大型河流流經。

基本信息
- 中文名
- 阿爾伯脫龍
- 外文名
- Alberta lizard/Lizard from Alberta
- 別名
- 亞伯達龍、艾伯塔龍
- 拉丁學名
- Albertosaurus
- 界
- 動物界
- 門
- 脊索動物門 Chordata
- 綱
- 蜥形綱Sauropsida
- 亞綱
- 目
- 蜥臀目Saurischia
- 亞目
- 獸腳亞目Theropoda
- 總目
- 恐龍總目
- 科
- 霸王龍科 Tyrannosauridae
- 亞科
- 阿爾伯塔龍亞科Albertosaurinae
- 屬
- 阿爾伯塔龍屬Albertosaurus
- 種
- 肉食阿爾伯塔龍A. sarcophagus
- 分布區域
- 加拿大阿爾伯塔省
- 超目
- 恐龍總目 Dinosauria
- 體征
- 頭部很大,成年艾伯塔龍的頭骨長約1.1米
- 生活時期
- 白堊紀