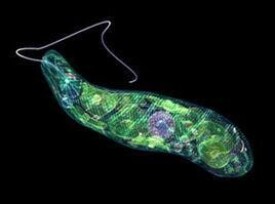
鞭毛綱(Mastigophora)物種約10000種。一般具鞭毛,以鞭毛為運動器,鞭毛通常有1~4條或稍多,鞭毛內周緣部排列9條雙聯體微管、中央有2條中央微管。無性繁殖一般為縱二分裂,有性繁殖為配子結合或整個個體結合。在環境不良的條件下一般能形成包囊。根據營養方式的不同,可分為兩個亞綱:植鞭亞綱Phytomastigina一般具有色素體能行光合作用,自由生活在淡水或海水中(自養),種類多,形狀各異,如眼蟲Euglena、盤藻Gonium、團藻Volvox、夜光蟲Noctiluca(腰鞭毛蟲類,可引發赤潮)、鐘罩蟲Dinobryon、尾窩蟲Uroglena、合尾滴蟲Synura。動鞭亞綱Zoomastigina無色素體,寄生、腐生或吞噬營養(異養),如利什曼原蟲Leishmania(黑熱病,白蛉子傳播),錐蟲Trypanosoma(昏睡病)、隱鞭蟲Cryptobia(寄生於魚鰓)、披髮蟲Ttrichonympha(共生於白蟻腸道,使其可以消化木材的纖維素)。
鞭毛綱是
原生動物門的一綱。以鞭毛運動。一般具有104根鞭毛,少數種類具有較多的鞭毛。有些種類具有色素體,能進行光合作用,自己製造食物,為植物性營養方式。有的通過體表滲透吸收周圍呈溶解狀態的物質,為腐生性營養。另一些則吞食固體的食物顆粒,為動物性營養方式。其中植物性營養方式為本綱所特有。有的種類根據生活環境條件的改變,或兼有兩種營養方式。在自然情況下都以二分裂法進行繁殖,除此,也有能形成
配子進行有性生殖的。
鞭毛蟲有自由生活於淡水或海水中的(眼蟲、夜光蟲等),也有寄生生活的。有不少寄生種類,能引起人或家畜的疾病(如引起黑熱病的利什曼原蟲,寄生於人或家畜體內的錐蟲、寄生於魚鰓的鰓隱鞭蟲等)。鞭毛綱還有些種類以致個或許多個體相聚,成為群體的(盤藻、團藻等)。
鞭毛綱(Mastigophora)動物在其成年階段都有一根、幾根或許多根鞭毛作為其運動細胞器,由於它們營養方式及生殖方式的原始性和多樣性,一般認為鞭毛蟲綱是最原始的原生動物,通過它可以將動物界與植物界聯繫起來。鞭毛蟲綱可以分為兩個亞綱:植鞭毛亞綱(Phytomastigina)及動鞭毛亞綱(Zoomastigina),前者都具有色素體,可行光合作用即進行植物性營養。澱粉及副澱粉是其主要的食物貯存物,有無性生殖和有性生殖,蟲體通常具有2根鞭毛,體表為
皮膜或纖維素細胞壁,自由生活。後者沒有色素體,行動物性營養或腐生性營養,
糖元是其食物貯存物,不行有性生殖,有一到數根鞭毛,蟲體表面只有細胞膜,除少數種類自由生活外,多數種類在多細胞動物體內營共生或寄生生活。
衣滴蟲 模式圖
1.以鞭毛為運動器,通常有1~4條,少數種類具有較多鞭毛
2.具有3種營養方式:植物性營養、動物性營養和滲透性營養
3.生殖方式多樣:無性生殖為縱二裂;有性生殖有同配生殖、異配生殖
4.環境不良時能形成包囊
綠眼蟲模式圖
鞭毛蟲類身體呈卵圓形、球形、長圓形等,通常能分出前後端,都以鞭毛作為運動器官,體表具細胞膜、或皮膜或
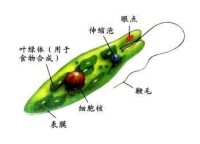
細胞壁。例如眼蟲(Euglena),體表具有皮膜,是由一層蛋白質組成螺旋形排列的條紋,在條紋上有嵴和溝,使皮膜呈現斜紋,眼蟲的皮膜很薄、富有彈性,使身體可以收縮改變形狀,做眼蟲式運動(euglenoid movement)。鞭毛蟲缺乏明顯的內質與外質的分化。眼蟲身體的前端有一凹陷的溝,稱儲蓄泡(reservoir)或稱胞咽,但它不是取食的器官,是水的貯存處及鞭毛著生部位,基體位於它的基部,由基體伸出兩根鞭毛,一根很長伸向體外,一根很短不伸出儲蓄泡。眼蟲和其他一些植鞭毛蟲類在身體的前端含有
類胡蘿蔔素(carotinoid)的脂類球集合而成的紅色眼點,這種色素球只有在基質中含有很少的氮及磷時才出現,否則將消失,它可以吸收光線。在鞭毛的基部具有副鞭毛體(paraflagellar body),是一種光敏感結構,當一側的光被眼點吸收后,可使它產生一個定向的光反應,所以副鞭毛體與眼點構成了某些植鞭毛蟲類的感光細胞器。眼蟲和許多淡水鞭毛蟲有一個或多個伸縮泡。眼蟲的伸縮泡位於身體前端,它是由幾個小泡收集了細胞質內過多的水分癒合而成,當伸縮泡充滿水分之後,排到儲蓄泡中,再通過其開口排到體外。某些鞭毛蟲沒有儲蓄泡,則伸縮泡內的水分通過體表排到體外。海洋或寄生的種類伸縮泡不發達或完全不存在。眼蟲及其他植鞭毛蟲類,細胞內散布有形狀、數目不等的色素體,眼蟲的色素體中含有
葉綠素a和b,胡蘿蔔素(carotin)和
葉黃素,使蟲體也表現出綠色。有些植鞭毛蟲的葉綠素被其他色素所隱蔽,而使鞭毛蟲表現出黃綠色或黃褐色等,與色素體常伴隨出現的是澱粉核(pyrenoid),它對副澱粉體的形成起作用。大多數鞭毛蟲具有一個大的細胞核,位於身體的近後端,少數種類具有2個核,核內有核內質(endosome)。眼蟲的細胞核在分裂間期時染色質濃縮,分裂過程中不行成
紡錘體,
核膜不消失,這是很原始的現象。
鞭毛蟲綱是以鞭毛作為運動器官,當鞭毛蟲有兩根或許多根鞭毛時,它們等長或不等長,如果不等長,其中一根起主導作用,另一根是附屬的,例如囊桿蟲(Peranema)。眼蟲在運動時也是一根鞭毛起作用。許多植鞭毛蟲的鞭毛可向一側或兩側伸出許多平行排列的細絲,稱茸鞭茸毛(mastigonemes),具有兩根不等長鞭毛的種類,一般主鞭毛是茸鞭茸毛,它的作用可能是逆轉水的推動力。一些寄生的鞭毛蟲其鞭毛不是遊離的,它由基體發生之後,與身體表面的細胞質膜之間有一原生質相連,在電子顯微鏡下觀察,是成行密集排列的絲狀結構連接在鞭毛與細胞之間,鞭毛的末端,離開身體之後是遊離的,例如錐蟲,這樣的結構有利於鞭毛蟲在粘滯性很高的血液或體液中運動。鞭毛運動的機制尚不十分清楚,不同的種類其鞭毛使用的方法並不完全相同。早期的理論認為鞭毛在運動時是螺旋旋轉,像
推進器一樣推動身體前進。現在知道鞭毛運動更多的是划動(rowing)及波動(undulating)。划動是鞭毛與附著點成直角方向在水中甩動,隨之是鞭毛的恢復打動。藉助於鞭毛甩動時水流的反作用力推動身體前進,並沿身體的縱軸向前旋轉前進,或左右擺動前進。波動運動是鞭毛在一個平面上呈波狀擺動。如果波狀擺動是由鞭毛的基部向端部發生,則驅使身體向相反的方向運動,即推動身體向後運動。如果波動是由鞭毛的端部向基部進行,則推動身體向前運動,在有兩個鞭毛的種類,運動的方向及路線是由主鞭毛決定的。關於鞭毛的結構與運動機制、人們普遍認為不是由於鞭毛中
微管纖維的收縮,因為它們在運動中並不改變長度,而是由成雙聯體的兩個亞纖維彼此滑動,引起鞭毛的彎曲,而亞纖維的滑動也像肌纖維的微絲滑動一樣,並且也是利用三磷酸腺苷(ATP)為能源。
鞭毛蟲類除了沒有化學合成營養物之外,可以行各種營養方式。植鞭毛蟲類均行植物性營養,也稱自養(autotrophs);動鞭毛蟲類行動物性或腐生性營養,或兩者兼行;統稱為異養(heterotrophs)。實際上對許多植鞭毛蟲並不能嚴格地區分自養與異養。例如眼蟲在光照條件下,它能自行合成有機物,如把眼蟲放在黑暗中飼養一段時間之後,眼蟲的葉綠素會逐漸消失,而改行腐生性營養,如果把眼蟲再放回光照條件下,它又可重新出現葉綠素,又恢復自養。一些腰鞭毛蟲類及金滴蟲類可以兼行自養與異養。有一種棕鞭毛蟲(Ochromonas)甚至可以兼行自養、動物性營養及腐生性營養三種營養方式。嚴格的異養僅出現在動鞭毛亞綱,它們或進行吞噬,或在鞭毛的基部形成胞咽取食。營養實驗研究表明,鞭毛蟲類比其他的原生動物的營養需求更簡單。植鞭毛蟲類生活的環境中只要有適當偽氮源及碳源,它就可以合成所需要的蛋白質;腐生性的鞭毛蟲要求環境中有氨鹽,就可以合成蛋白質;在營動物性營養的鞭毛蟲則要求環境中有
氨基酸及
蛋白腖以合成所需要的蛋白質。其他綱原生動物則需要現成的食物(蛋白質),其自行合成食物的生理機能已經消失。植鞭毛蟲類食物的貯存形式是植物澱粉,副澱粉體、脂肪及麥清蛋白(leucosin),動鞭毛蟲類食物的貯存形式是糖元。
鞭毛蟲類的無性生殖主要是
二分裂,其中多數又是縱二分裂,例如眼蟲,細胞分裂由中心粒(centrioles)或基體的分裂開始,由每個中心粒產生一個新的基體及一根鞭毛,然後每一個新鞭毛與一個老鞭毛根部癒合,同時核也進行
有絲分裂,但核膜不消失、不行成紡錘體,隨後
細胞質由前向後分裂,並複製新的
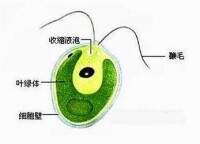
細胞器,最後形成兩個相似的子細胞。除了縱分裂,有些種類生殖時可以是:斜分裂或橫裂,例如角藻;或多分裂,例如某些腰鞭毛蟲類及錐蟲等;也可以出現出芽生殖,例如夜光蟲(Noctiluca),即由母體上可以長出許多小的芽體。眼蟲分裂時一個子細胞保留舊鞭毛,一個子細胞再複製一個新鞭毛,但也有的種原來的鞭毛完全消失,兩個子細胞均產生新的鞭毛;如果蟲體有幾根鞭毛,分裂前部分鞭毛已經複製,分裂時再均等的分配在兩個子細胞中。在多鞭毛的種,分裂時鞭毛均等的分配在兩個子細胞中,然後每個子細胞再複製一定數量的鞭毛使之恢復到應有的鞭毛數。腰鞭毛蟲類體表常具有甲板(armored),分裂時甲板或消失,或均等或不均等的分配到子細胞中。角藻常進行不完全的分裂,結果使多次分裂的子體連成一個長鏈。群體生活的種類如團藻(Volvox)分裂的結果在群體中形成子群體。總之,無性生殖的方式因種而不同。有性生殖主要出現在植鞭毛蟲類及複雜的多鞭毛蟲類。它們或是同配生殖或異配生殖,例如衣滴蟲(Chlamydomonas)生殖時兩個蟲體彼此融合,所以蟲體本身又是配子,減數分裂發生在配子融合之後,衣滴蟲是同配生殖的典型例子。實球蟲(Pandorina)是異配生殖,融合的兩個配子僅略有大小的區別,大配子仍有鞭毛。團藻是以典型的異配生殖形成精子與卵子,其精子與卵子可能來自同一個群體(雌雄同群體),也可能來自不同的群體(雌雄異群體),卵直接來自群體的生殖細胞其體積增大、沒有鞭毛、不會運動。精子是由生殖細胞經多次分裂形成,精子成束、具鞭毛、能遊動。卵受精之後,外面可分泌厚壁形成卵囊以越冬。鞭毛蟲類的包囊形成是很普遍的,特別是營動物性營養的種類、具葉綠素的種類較少發現包囊。
植鞭毛亞綱 Phytomastigina
┗金滴蟲目 Chrysomonadid
┗隱滴蟲目Cryptomonadida
┗腰鞭毛目 Dinoflagellida
┗眼蟲目 Euglenida
┗綠滴蟲目 Chloromonadida
┗團藻目 Volvocida
動鞭毛亞綱 Zoomastigina
┗領鞭毛目 Choanoflagellida
┗根足鞭毛蟲目Rhizomastigida
┗動質體目 Kinetoplastida
┗曲滴蟲目 Retortamonadida
┗雙滴蟲目 Diplomonadida
┗毛滴蟲目 Trichomonadida
┗超鞭毛目 Hypermastigida
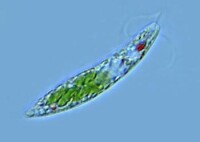
綠眼蟲 Euglena viridis
眼蟲是眼蟲目Euglenida眼蟲屬Euglena生物的統稱,在植物學中稱裸藻,是一類介於動物和植物之間的單細胞
真核生物。淡水中習見的眼蟲有:綠眼蟲Euglena viridis,體紡錘形,前端鈍圓,後端寬,末端尖呈尾狀。鞭毛與體等長,色素體1個,星狀。梭眼蟲Euglena acus,長紡錘形,鞭毛短,色素體多個。長眼蟲Euglena deses,體圓柱形,狹長,鞭毛約為體長的1/3~1/2。螺紋眼蟲Euglena spirogyra,體易變形,體表螺旋形帶紋明顯,鞭毛短。扁眼蟲Phacus,體呈寬卵圓形,背腹扁,後端尖刺狀,鞭毛與體等長。眼蟲生活在有機物質豐富的水溝、池沼或緩流中。但在河堤、海灣濕土或含鹽沼澤中亦有之,此外在其它藻類體上、植物碎片、及小甲殼類的體上亦能見到。至於營有機性的種類則多見之於下水道的水內。溫暖季節可大量繁殖常使水呈綠色。眼蟲體呈綠色梭形,長約60μm,前端鈍圓,後端尖。在蟲體中部稍後有一個大而圓的核,生活時是透明的。體表覆以具彈性的、帶斜紋的表膜。經電子顯微鏡研究,表膜即質膜或稱三分質膜。表膜是由許多螺旋狀的條紋聯結而成,每一個表膜條紋的一邊有向內的溝(groove),另一邊有向外的嵴(crest)。一個條紋的溝與其鄰接條紋的嵴相關聯(似關節)。眼蟲生活時,表膜條紋彼此相對移動,可能是由於嵴在溝中滑動的結果。表膜下的粘液體(mucusbody)外包以膜,與體表膜相連續,有粘液管通到嵴和溝。粘液對溝嵴聯結的“關節”可能有滑潤作用。表膜覆蓋整個體表、胞咽、儲蓄泡、鞭毛等。使眼蟲保持一定形狀,又能作收縮變形運動。眼蟲體前端有一胞口(cytostome).向後連一膨大的儲蓄泡(reservoir),從胞口中伸出一條鞭毛(flagellum)。鞭毛是能動的細胞表面的突起。鞭毛下連有2條細的軸絲(axoneme)。每一軸絲在儲蓄泡底都和一基體(basal body)相連,由它產生出鞭毛。基體對蟲體分裂起著中心粒的作用。從一個基體連一細絲(根絲體rhizoplast)至核,這表明鞭毛受核的控制。鞭毛最外為細胞膜,其內由縱行排列的微管(microtubule)組成。周圍有9對聯合的微管(雙聯體doublets), 中央有2個微管。每個雙聯體上有2個短臂(arms),對著下一個雙聯體,各雙聯體有放射輻(radial spokeS)伸向中心。在雙聯體之間又有具彈性的連絲(links)。微管由微管蛋白(tubulin)組成,微管上的臂是由動力蛋白(dynein)組成,具有
ATP酶的活性。實驗證明,鞭毛的彎曲,是由於雙聯體微合彼此相對滑動的結果,如圖所示,在彎曲的內、外側放射輻的間隔不改變,彎曲是由於彎曲的外側微管和放射輻對於彎曲內側的微管和放射輻的相對滑動。一般認為臂能使微管滑動(很像肌肉收縮時,橫橋在粗、細肌絲間的滑動),臂上的ATP酶分解ATP提供能量。眼蟲借鞭毛的擺動進行運動。眼蟲在運動中有趨光性,這是因為在鞭毛基都緊貼著儲蓄泡有一紅色眼點(stigma),靠近眼點近鞭毛基都有一膨大部分,能接受光線,稱光感受器(photoreceptor)。眼點是由埋在無色基質中的類胡蘿蔔素(carotenoid)的小顆粒組成的。也有人認為是由胡蘿蔔素(carotene)組成的,或是由β-紅蘿蔔素與
血紅素組成的。眼點呈淺杯狀,光線只能從杯的開口面射到光感受器上,因此,眼蟲必須隨時調整運動方向,趨向適宜的光線。現在有些學者認為,眼點是吸收光的“遮光物”(light absorbing shade),在眼點處於光源和光感受器之間時,眼點遮住了光感受器,並切斷了能量的供應,於是在蟲體內又形成另一種調節,使鞭毛打動,調整蟲體運動,讓光線的連續地照到光感受器上。這樣連續調節使眼蟲趨向光線。眼點和光感受器普遍存在於綠色鞭毛蟲,這與它們進行光合作用的營養方式有關。在眼蟲的細胞質內有葉綠體(chloroplast)。葉綠體的形狀(如卵圓形、盤狀、片狀、帶狀、星狀等)、大小、數量及其結構(有無蛋白核及副澱粉鞘)為眼蟲屬、種的分類特徵。眼蟲主要通過葉綠素在有光的條件下利用光能進行光合作用,把二氧化碳和水合成糖類,這種營養方式(與一般綠色植物相同),稱為光合營養(phototrophy)。製造的過多食物形成一些半透明的副澱粉粒(Paramylum granule)儲存在細胞質中。副澱粉粒與澱粉相似,是糖類的一種,但與碘作用不呈藍紫色。副澱粉粒是眼蟲類特徵之一,其形狀大小也是其分類的依據。在無光的條件下,眼蟲也可通過體表吸收溶解於水中的有機物質。這種營養方式稱為滲透營養(osmotrophy)。眼蟲前端的胞口是否取食固體食物顆粒還有異議。但是已肯定經過胞口可以排出體內過多的水分。在儲蓄泡旁邊有一個大的伸縮泡(contractile vacuole),它的主要功能是調節水分平衡。收集細胞質中過多的水分(其中也有溶解的代謝廢物)。排入儲蓄泡,再經胞口排出體外。眼蟲在有光的條件下,利用光合作用所放出的氧進行呼吸(氧化)作用,呼吸作用所產生的二氧化碳,又被利用來進行光合作用。在無光的條件下,通過體表吸收水中的氧,排出二氧化碳。眼蟲的生殖方法一般是縱二分裂,這也是鞭毛蟲綱的特徵之一。先是核進行有絲分裂,在分裂時核膜不消失,基體複製為二,繼之蟲體開始從前端分裂,鞭毛脫去,同時由基體再長出新的鞭毛,或是一個保存原有的鞭毛,另一個產生新的鞭毛。胞口也縱裂為二,然後繼續由前向後分裂,斷開成為2個個體。在環境不良的條件下。如水池乾涸.眼蟲體變圓,分泌一種膠質形成包囊,將自己包圍起來。剛形成的包囊、可見有眼點,綠色,以後逐流變為黃色,眼點消失,代謝降低,可以生活很久,隨風散佈於各處。當環境適合時,蟲體破囊而出,在出囊前進行一次或幾次縱分裂。包囊形成對眼蟲度過不良環境是一種很好的適應。多年來用眼蟲進行基礎理論的研究取得不少成果。不僅對遺傳變異理論的探討有意義,而且對了解有色、無色鞭毛蟲類動物間的親緣關係,對了解動、植物的親緣關係都有重要意義。近年來也有用眼蟲作為有機物污染環境的生物指標,用以確定有機污染的程度,另外眼蟲對凈化水的放射性物質也有作用。
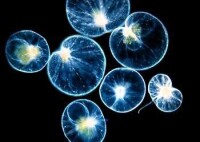
夜光蟲 Noctiluca
夜光蟲Noctiluca屬腰鞭毛目Dinoflagellida。它們在夜間由於海水波動的刺激能發光,因而得名。夜光蟲的身體為圓球形,直徑為1毫米左右,顏色發紅,細胞質密集於球體的一部分,其內有核,其他部分由細胞質放散成粗網狀,在網眼間充滿液體。有兩根鞭毛,一根較大,稱為觸手,另一根較小。它的繁殖為分裂法和出芽法兩種,後者在身體表面生出很多小的個體,脫離母體后發育成新的個體。例如閃光夜光蟲身體的直徑為0.5~2毫米,肉眼看到的是一個個晶亮的小球,有透明的細胞膜、網狀分散的細胞質、濃密的細胞核、一根細小的鞭毛以及原生質突起形成的粗大可動的觸手。因其體內含有許多擬脂顆粒,故受到機械刺激時能發光。在海水中生活的夜光蟲和其他一些腰鞭毛蟲(如溝腰鞭蟲Gonyaulax 、裸甲腰鞭蟲Gymnodinium等)大量繁殖,每立方米海水可多達20~40百萬個個體,致使海水呈現紅色或褐色,稱為
赤潮,由於它們排出大量代謝產物以及腐敗海水,造成沿海魚類及養殖貝類的大量死亡。腰鞭毛目Dinoflagellida,具兩根鞭毛,分別位於身體中部的橫溝及後部的縱溝內,橫溝內的鞭毛使身體旋轉、縱溝內鞭毛推動身體前進。色素體黃色或褐色,少數種無色素體。一些種類體表裸露,如Gymnodinium,多數種類體表有纖維素形成的薄膜、或甲板,如薄甲藻Glenodinium、膝溝藻Gonyaulax等。眼點通常是存在的。無性生殖有的種可行橫裂及斜裂,如角藻Ceratium。
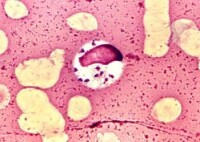
杜氏利什曼原蟲
利什曼原蟲Leishmania屬動質體目Kinetoplastida(具1~2根鞭毛,所有的成員都有一動質體,常與根絲體相連,其中含有DNA,位於大的延伸的
線粒體內,身體一側有波動膜)。生活史有前鞭毛體(promastigote)和無鞭毛體(amastigote)兩個時期。前者寄生於
節肢動物(
白蛉)的消化道內,後者寄生於哺乳動物或爬行動物的細胞內,通過白蛉傳播。對人和哺乳動物致病的利什曼原蟲有:引起人體內臟利什曼病的為
杜氏利什曼原蟲Leishmania donovani,引起皮膚利什曼病的為熱帶利什曼原蟲Leishmania tropica和墨西哥利什曼原蟲Leishmania mexicana,引起粘膜皮膚利什曼病的為巴西利什曼原蟲Leishmania araziliensis等。我國的黑熱病是由杜氏利什曼原蟲引起的。杜氏利什曼原蟲的無鞭毛體主要寄生在肝、脾、骨髓、淋巴結等器官的
巨噬細胞內,常引起全身癥狀,如發熱、
肝脾腫大、貧血、鼻衄等。在印度,患者皮膚上常有暗的色素沉著,並有發熱,故又稱Kala-azar,即黑熱的意思。因其致病力較強很少能夠自愈,如不治療常因併發病而死亡。解放前,全國黑熱病患者較多。主要流行於長江以北廣大地區,為我國五大寄生蟲病之一。解放后在黨的領導下,在各流行區建立專門的防治機構,發動群眾從治病、消滅病犬和白嶺子三方面進行防治。現已在全國範圍內基本上控制了黑熱病的流行。
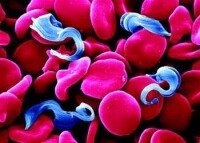
錐體蟲 Trypanosoma
錐體蟲Trypanosoma動質體目錐蟲科錐蟲屬。體呈柳葉狀,運動胞器是唯一的一根鞭毛。寄生在血液中的鞭毛蟲,其鞭毛與蟲體之間連成為波動膜,藉以增強在粘滯性較高的血液中的活動能力。
細胞核 1個。體內有一稱作動基體的特殊細胞器,位於錐蟲體內唯一的線粒體內,含有豐富的DNA,謂之動基體DNA(KDNA),能自我複製。動基體的確切功能至今不明。繁殖方式為二分裂。種類多,分別寄生在各種脊椎動物(魚類、兩棲類、爬行類、鳥類和哺乳類)的血液和
組織液中。有個別種類如枯氏錐蟲則寄生在人的細胞內。除馬媾疫錐蟲是通過交媾直接傳播外,所有寄生在脊椎動物的錐蟲均要依賴某些昆蟲(如
采采蠅等)進行傳播。帶有錐蟲的脊椎動物血液被媒介動物吸食后,錐蟲因生活環境改變,在媒介動物消化道中依次經過無鞭毛體或前鞭毛體上鞭毛體和后循環錐鞭毛體幾個不同的發育階段。在媒介動物中只有這種后循環錐蟲對宿主才有感染力。當媒介再次叮咬宿主時,便把后循環錐蟲輸入宿主體內。伊氏錐蟲在媒介消化道中不進行任何發育,它靠媒介在吸血后短時間內作機械傳播。錐蟲主要通過胞飲作用從宿主獲得營養。所有錐蟲均可在體外連續培養。錐蟲是人和家畜重要的寄生蟲之一。按其傳播方式可分為兩大類:①糞便型,通過被后循環錐蟲污染的糞便傳播;②唾液型,后循環錐蟲經
唾液腺傳播。對人有嚴重致病作用的錐蟲有:羅得西亞錐蟲、甘比亞錐蟲、枯氏錐蟲等。前兩者主要流行於非洲各地,引起所謂非洲睡眠病;後者主要分佈在南美洲(特別是巴西),引起美洲錐蟲病,即夏格氏病。中國至今還沒有發現人體錐蟲的病例。對家畜有嚴重致病作用的錐蟲有布氏錐蟲、活潑錐蟲、剛果錐蟲、伊氏錐蟲和馬媾疫錐蟲等。前三者主要流行於非洲,中國尚無記載。伊氏錐蟲在中國分佈甚廣,牛、馬伊氏錐蟲病即所謂蘇拉病,在中國南方各省流行相當嚴重,造成大批家畜死亡。由於錐蟲存在抗原變異的特性,至今仍未獲得有效的疫苗。寄生家畜的布氏錐蟲和寄生鼠的路氏錐蟲是實驗室常用的研究材料。