細胞分化
細胞分化
細胞分化(cell differentiation)是指同一來源的細胞逐漸產生出形態結構、功能特徵各不相同的細胞類群的過程,其結果是在空間上細胞產生差異,在時間上同一細胞與其從前的狀態有所不同。細胞分化的本質是基因組在時間和空間上的選擇性表達,通過不同基因表達的開啟或關閉,最終產生標誌性蛋白質。一般情況下,細胞分化過程是不可逆的。然而,在某些條件下,分化了的細胞也不穩定,其基因表達模式也可以發生可逆性變化,又回到其未分化狀態,這一過程稱為去分化(dedifferentiation)。
細胞分化
細胞分化
在細胞分化中,細胞核起決定作用。一般認為細胞核內含有該種生物的全套遺傳信息。在條件具備時,它可使所在細胞發育分化為由各種類型細胞所組成的完整個體。從培養的煙草,髓部小塊形成的組織團塊上取脫落的細胞,單個分離培養能得到有根和葉的幼芽,再移植到土壤中,會長出開花的植物。在兩棲類,把囊胚期和早期原腸胚的細胞核移植到事先已經去掉細胞核的卵內能使卵正常發育,說明動植物體細胞的核是全能的。
分化與細胞質之間的關係可以從卵質談起。如馬副蛔蟲受精后,所有經過染色體消減的細胞都發育為體細胞(見生殖質)。許多動物卵子細胞質的分佈有明顯的區域性。這種區域性雖然不影響染色體的行為,但對於以後胚胎器官發育卻有決定性作用。中國胚胎學家童第周等利用核移植的技術,也證實了卵質在性狀發生中的作用。他們把金魚囊胚期細胞核移到去核的鰟鮍魚卵子中;雖然發育到幼魚的例子極少,但是發育的過程都比較正常,一些基本的發育的特點,如胚胎的背腹性,對稱性以及早期的卵裂進程等都和鰟鮍魚一樣,幼魚的體形也和鰟鮍魚的幼魚沒有區別。這些性狀的出現似乎完全根據細胞質。細胞質對細胞核的作用,還表現在對核功能活動的影響。如培養的人宮頸上皮癌細胞——HeLa細胞——的DNA和RNA合成都很活躍;雞的紅細胞雖然有核,但是處於不活躍狀態,不進行DNA合成,RNA合成也很微弱。用細胞融合的方法,使去掉細胞核的HeLa細胞的細胞質和雞的紅細胞融合,便可使後者的細胞核體積增大,濃縮的染色質變得鬆散,原來已經失去的合成RNA和DNA的功能在寄主HeLa細胞質的影響下,重新恢復了。
卵母細胞的細胞核並不位於細胞中央,而是在細胞外周靠近表面的地方,極體就是從這裡形成並釋放出來的,通常把極體釋放的位點稱為動物極,而相對的一極稱為植物極。
動物卵細胞中,貯存有2萬-5萬種不同核苷酸序列的mRNA,專供受精卵的啟動、分化和發育之用。卵細胞中的mRNA並非均勻分佈的,而是位於特定的空間。因此,卵細胞質的特性決定了子細胞核的分化命運,如昆蟲是以表面卵裂的方式形成胚層細胞的。
細胞間的相互作用是各式各樣的,可以是誘導作用,也可以是抑制作用。就作用方式來說,有的作用需要細胞的直接接觸,另一些所需要的可能是間隔一定距離的化學物質的擴散。
(1)胚胎誘導:在胚胎髮育過程中,一部分細胞影響相鄰細胞向一定方向分化的作用稱為胚胎誘導,如中胚層形成的脊索可誘導其頂部的外胚層發育成神經板、神經溝和神經管,視細胞可誘導其外面的外胚層形成晶體,而晶體又可誘導外胚層形成角膜。
誘導不但在中軸器官的形成中起作用,也在以後器官的發生中起作用。例如間質細胞的存在對體內腺體上皮的形成和分化是必不可少的。這些腺體包括甲狀腺、胸腺、唾腺和胰腺,它們對間質細胞的依賴程度有很大差異。在離體條件下,胰腺原基只要有間質細胞存在就可以繼續發育。
(2)分化抑制:用含有成蛙心組織的培養液培養蛙胚,則蛙胚不能發育出正常的心臟,若去除成蛙心組織,則蛙胚發育正常,這說明分化成熟的細胞可以產生某種物質,抑制相鄰細胞發生同樣的分化,這種作用稱為分化抑制。
如在蠑螈幼蟲或成體摘除水晶體后,可以從背部的虹彩再生出一個新的。進一步的分析指出,再生水晶體的能力局限在虹彩背部的邊緣層。如把這部分組織移到另一個摘除水晶體的眼睛,不是位於背部,而是使它位於腹部,仍舊可以由它再生出水晶體。
既然這部分細胞有生長水晶體的能力,為什麼在正常的眼睛里不表現?如把虹彩的背部移到另一隻未摘除水晶體的眼睛里,不管使它位於那一部位,都長不出水晶體。如在摘除水晶體的眼睛里,經常注射完整的(帶有水晶體的)眼腔液體,在注射期間,虹彩背部的細胞也長不出水晶體。由此可見,虹彩背部的細胞本來具有產生水晶體的能力,正常水晶體會產生一種物質,對此起抑制作用。
(3)細胞數量效應:當小鼠胚胎胰腺原基在體外進行組織培養時,可發育成具有功能的胰腺組織,但是,如果把胰腺原基切成8小塊分別培養,則都不能形成胰腺組織;如果再把分開的小塊合起來,又可形成胰腺組織。可見細胞數量對誘導組織形成可能也是必要的。
(4)細胞外基質:細胞外基質在胚胎髮育和細胞分化中也具有重要的作用。
(5)激素的作用:激素對細胞分化的影響可看做是遠距離細胞間的相互作用,如昆蟲的保幼激素和蛻皮激素。前者的功能是保持幼蟲特徵,促進成蟲器官原基的發育,後者的功能是促進蛻皮和成蟲形成的出現。當兩者保持一定的比例時,幼蟲化蛹,變為成蟲。成蟲期又開始合成保幼激素,促進性腺的發育。
細胞間的相互作用是各式各樣的,可以是誘導作用,也可以是抑制作用。就作用方式來說,有的作用需要細胞的直接接觸,另一些所需要的可能是間隔一定距離的化學物質的擴散。
①誘導作用。兩棲類胚胎背部的外胚層細胞,在脊索中胚層的作用下,分化為神經細胞,以後發育為神經系統。這種中軸器官的誘導作用在脊椎動物具有普遍性,一般認為,脊索中胚層細胞釋放某種物質,誘導外胚層細胞分化為神經組織。誘導不但在中軸器官的形成中起作用,也在以後器官的發生中起作用。例如間質細胞的存在對體內腺體上皮的形成和分化是必不可少的。這些腺體包括甲狀腺、胸腺、唾腺和胰腺,它們對間質細胞的依賴程度有很大差異。在離體條件下,胰腺原基只要有間質細胞存在就可以繼續發育。
②抑制作用。如在蠑螈幼蟲或成體摘除水晶體后,可以從背部的虹彩再生出一個新的。進一步的分析指出,再生水晶體的能力局限在虹彩背部的邊緣層。如把這部分組織移到另一個摘除水晶體的眼睛,不是位於背部,而是使它位於腹部,仍舊可以由它再生出水晶體。
既然這部分細胞有生長水晶體的能力,為什麼在正常的眼睛里不表現?如把虹彩的背部移到另一隻未摘除水晶體的眼睛里,不管使它位於那一部位,都長不出水晶體。如在摘除水晶體的眼睛里,經常注射完整的(帶有水晶體的)眼腔液體,在注射期間,虹彩背部的細胞也長不出水晶體。由此可見,虹彩背部的細胞本來具有產生水晶體的能力,正常水晶體會產生一種物質,對此起抑制作用。
細胞分化中基因表達的調節控制是一個十分複雜的過程,在蛋白質合成的各個水平,從mRNA的轉錄、加工到翻譯,都會有調控的機制。在DNA水平也存在調控機制(如基因的丟失、放大、移位重組、修篩以及染色質結構的變化等)。不同的細胞在其發育中的基因表達的調節控制不同;相同的細胞在其發育的各階段中,調節控制的機制不同。
一、全能性、多能性和單能性受精卵能夠分化出各種細胞、組織,形成一個完整的個體,所以把受精卵的分化潛能稱為全能性。隨著分化發育的進程,細胞逐漸喪失其分化潛能。從全能性到多能性,再到單能性,最後失去分化潛能成為成熟定型的細胞。植物的枝、葉、根都有可能長成一株完整的植株,細胞培養的結果也證明即使高度分化的植物細胞也可以培養成一個完整的植株,因此可以說絕多數植物細胞具有全能性。成熟動物細胞顯然不具備全能性。其原因並非在細胞核而在細胞質,如大量的核移殖實驗證實,分化細胞的核仍保留完整的基因組DNA。中國發育生物學家童第周1978年成功地將黑斑蛙成熟的細胞核移入去核的受精卵細胞內,培育出了蝌蚪。60年代的爪蟾和80年代小鼠的核移殖,90年代末多利羊的誕生都證明了分化細胞具有完整的基因組DNA。在人的一生中,皮膚、小腸和血液等組織需要不斷地更新,這個任務是由幹細胞完成的。幹細胞是一類具有分裂和分化能力的細胞,多能幹細胞可以分化出多種類型的細胞,但它不可能分化出足以構成完整個體的所有細胞,所以多能幹細胞的分化潛能稱為多能性(pluripotent)。單能幹細胞來源於多能幹細胞,具有向特定細胞系分化的能力,也稱為祖細胞(progenitor)。二、幹細胞的特點幹細胞具有以下生物學特點:①終生保持未分化或低分化特徵;②在機體的中的數目、位置相對恆定;③具有自我更新能力;④能無限制的分裂增殖;⑤具有多向分化潛能,能分化成不同類型的組織細胞,造血幹細胞、骨髓間充質幹細胞、神經幹細胞等成體幹細胞具有一定的跨系、甚至跨胚層分化的潛能;⑥分裂的慢周期性,絕大多數幹細胞處於G0期;⑦通過兩種方式分裂,對成分裂和不對稱分裂前者形成兩個相同的幹細胞,後者形成一個幹細胞和一個祖細胞。根據幹細胞的分化能力,可以分為全能幹細胞、多能幹細胞和單能幹細胞。全能幹細胞可以分化為機體內的任何一種細胞,直至形成一個複雜的有機體。多能幹細胞可以分化為多種類型的細胞,如造血幹細胞可以分化為12種血細胞。有些文獻中將分化潛能更廣的細胞叫做多潛能幹細胞(pluripotentstemcell),如骨髓間充質幹細胞,而把向某一組織類型細胞分化的幹細胞叫做多能幹細胞(multipotentstemcell),如前面提到的造血幹細胞。單能幹細胞只能分化為一種類型的細胞,而且自我更新能力有限。三、胚胎幹細胞根據個體發育過程中出現的先後次序不同,幹細胞又可分為胚胎幹細胞和成體幹細胞。胚胎幹細胞(embryonicstemcells,ESC)是指從胚胎內細胞團或原始生殖細胞篩選分離出的具有多能性或全能性的細胞,此外也可以通過體細胞核移植技術獲得。ESC能表達POU家族的轉錄因子Oct-3/4;在移植后能形成的畸胎瘤,在體外適當條件下能分化為代表三胚層結構的體細胞ESC的用途主要有:①克隆動物,由體細胞作為核供體進行克隆動物生產,雖然易於取材,但克隆動物個體中表現出嚴重的生理或免疫缺陷,而且多為致命性的;②轉基因動物,以ESC細胞作為載體,可大大加快轉基因動物生產的速度,提高成功率;③組織工程,人工誘導ESC定向分化,培育出特定的組織和器官,用於醫學治療的目的。
四、再生狹義地講再生指生物的器官損傷后,剩餘的部分長出與原來形態功能相同的結構的現象稱為再生,如壁虎的尾、蠑螈的肢、螃蟹的足,在失去后又可重新形成,海參可以形成全部內臟,水螅、蚯蚓、蝸蟲等低等動物的每一段都可以形成一個完整的個體等等。但是從廣義的角度來看再生是生命的普遍現象,從分子、細胞到組織器官都具有再生現象。再生的形式:生理性再生:即細胞更新,如人體內每秒中約有600萬個新生的紅細胞替代相同數量死亡的紅細胞。修復性再生:許多無脊椎動物用這種方式來形成失去的器官,如上述提到的壁虎的尾和螃蟹的肢。重建:是人工實驗條件下的特殊現象。如人為將水螅的一片組織分散成單個細胞。在懸液中,這些細胞重新聚集,在幾天至幾周以後,形成一條新的水螅。
腫瘤患者經常會聽到類似“高分化鱗癌”,“低分化腺癌”等專業術語。大多病人在患病之初往往不了解這些術語的意義。現在我們就來介紹一個關於癌細胞非常關鍵的一個術語--腫瘤細胞分化程度(Differentiation)。
所謂分化程度就是指腫瘤細胞接近於正常細胞的程度。分化得越好(稱為“高分化”就意味著腫瘤細胞越接近相應的正常發源組織;而分化較低的細胞(稱為“低分化”或“未分化”)和相應的正常發源組織區別就越大,腫瘤的惡性程度也相對較大。
按照腫瘤分化的程度,病理專家通常將其分為三個病理等級,並用英文字母G(代表Grade,即分化)來表示。級別越高表示細胞分化程度越差。
G1,即高分化,細胞分化程度較好。一般來說,G1的腫瘤細胞分裂速度較慢。
G2,即中分化,細胞分化程度居中。
G3,即低分化,細胞分化程度較差。腫瘤細胞分裂速度較快。
可以說腫瘤細胞的分化程度越差,它的惡性程度就越高,腫瘤體生長較迅速,而且容易發生轉移。分化好的腫瘤一般生長較慢,而且在治療后不易複發。但是,對不同腫瘤來說,腫瘤細胞的分化程度和病人的預后並不一定都有直接關係。從治療的角度上來說,某些分化程度低的細胞對於化療和放療更敏感,換言之,這些分化程度越低的腫瘤越容易通過化放療來治療。因此,並非高分化腫瘤的預后都好於低分化腫瘤。比如常見的血液惡疾淋巴癌,某些中高分化的淋巴癌通過化療和放療的聯合治療方法,治癒率可達40%左右。而大多的慢性淋巴癌(屬低分化),病情的發展往往非常緩慢,可持續幾年甚至十幾年,但藥物治療對慢性淋巴癌卻幾乎沒有治癒的效果。鼻咽癌的診治中也有類似的情況。又如口腔或咽喉部鱗癌,腫瘤細胞的分化程度和病人的預后則沒有直接關聯。
總之,對於不同的腫瘤來說,細胞的分化程度有著不同的意義。腫瘤細胞的分化程度是癌症診斷和治療中一個重要的參考的數據,但治療的效果,還是需要結合癌症的種類,分期,治療方法來綜合判斷。
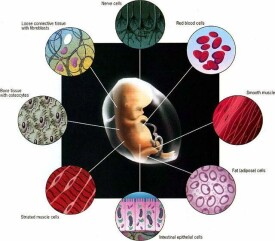
基本信息
- 中文名
- 細胞分化
- 外文名
- cellular differentiation
- 應用學科1
- 動物學-動物胚胎學
- 分化特點
- 持久、穩定、不可逆、普遍
- 核心問題
- 基因調控
- 應用學科2
- 胚胎學-胚胎概論