脂肪酸氧化
脂肪酸氧化
脂肪酸氧化,fatty acid oxidation,是指油脂水解產生的甘油和脂肪酸在供氧充足的條件下,可氧化分解生成二氧化碳和水,並釋放出大量能量供機體利用,在體內脂肪酸氧化以肝和肌肉最為活躍,而在神經組織中極為低下。脂肪酸氧化的方式有β-氧化和特殊氧化方式。特殊氧化方式有:丙酸氧化、α-氧化、ω-氧化、不飽和脂肪酸氧化。
中文名稱:脂肪酸氧化
英文名稱:fatty acid oxidation
脂肪酸氧化的方式有β-氧化和特殊氧化方式。特殊氧化方式有:丙酸氧化、α-氧化、ω-氧化、不飽和脂肪酸氧化。
過程可概括為活化、轉移、β氧化及最後經三羧酸循環被徹底氧化生成CO2和H2O並釋放能量幾階段。
(1)脂肪酸的活化:脂肪酸的氧化首先須被活化,在ATP、CoA-SH、Mg2+存在下,脂肪酸由位於內質網及線粒體外膜的脂醯CoA合成酶催化生成脂醯CoA。活化的脂肪酸不僅為一高能化合物,而且水溶性增強,因此提高了代謝活性。
(2)脂醯CoA的轉移:脂肪酸活化是在胞液中進行的,而催化脂肪酸氧化的酶系又存在於線粒體基質內,故活化的脂醯CoA必須先進入線粒體才能氧化,但已知長鏈脂醯輔酶A是不能直接透過線粒體內膜的,因此活化的脂醯CoA要藉助L-肉鹼(L-camitine),即L-3-羥基-4-三甲基銨丁酸,而被轉運入線粒體內,在線粒體內膜的外側及內側分別有肉鹼脂醯轉移酶I和酶Ⅱ,兩者為同工酶。位於內膜外側的酶Ⅰ,促進脂醯CoA轉化為脂醯肉鹼,後者可藉助線粒體內膜上的轉位酶(或載體),轉運到內膜內側,然後,在酶Ⅱ催化下脂醯肉鹼釋放肉鹼,后又轉變為脂醯CoA.這樣原本位於胞液的脂醯CoA穿過線粒體內膜進入基質而被氧化分解。一般10個碳原子以下的活化脂肪酸不需經此途徑轉運,而直接通過線粒體內膜進行氧化。
(3)脂醯CoA的β氧化:脂醯CoA進入線粒體基質后,在脂肪酸β氧化酶系催化下,進行脫氫、加水,再脫氫及硫解4步連續反應,最後使脂醯基斷裂生成一分子乙醯CoA和一分子比原來少了兩個碳原子的脂醯CoA.因反應均在脂醯CoA烴鏈的α,β碳原子間進行,最後β碳被氧化成醯基,故稱為β氧化。
(4)硫解:在β-酮脂醯CoA硫解酶(β-ketoacyl-CoA thiolase)作用下,β-酮脂醯CoA被一分子CoA所分解,生成一分子乙醯CoA和一分子比原來少兩個碳原子的脂醯CoA。少了兩個碳原子的脂醯CoA,可再次進行脫氫、水化、再脫氫和硫解反應,每經歷上述幾步后即脫下一個二碳單位(乙醯CoA)。
β-氧化
概述
脂肪酸的β-氧化 肝和肌肉是進行脂肪酸氧化最活躍的組織,其最主要的氧化形式是β-氧化。此過程可分為活化,轉移,β-氧化共三個階段。
在肝臟中,脂肪酸經β-氧化作用生成乙醯輔酶A。2分子乙醯輔酶A可縮合生成乙醯乙酸。乙醯乙酸可脫羧生成丙酮,也可還原生成β-羥丁酸。乙醯乙酸、β-羥丁酸和丙酮總稱為酮體。
β-氧化前提
1>脂肪酸的活化
和葡萄糖一樣,脂肪酸參加代謝前也先要活化。其活化形式是硫酯——脂肪醯CoA,催化脂肪酸活化的酶是脂醯CoA合成酶(acyl CoA synthetase)。
活化後生成的脂醯CoA極性增強,易溶於水;分子中有高能鍵、性質活潑;是酶的特異底物,與酶的親和力大,因此更容易參加反應。
脂醯CoA合成酶又稱硫激酶,分佈在胞漿中、線粒體膜和內質網膜上。胞漿中的硫激酶催化中短鏈脂肪酸活化;內質網膜上的酶活化長鏈脂肪酸,生成脂醯CoA,然後進入內質網用於甘油三酯合成;而線粒體膜上的酶活化的長鏈脂醯CoA,進入線粒體進入β-氧化。
2>脂醯CoA進入線粒體
長鏈脂肪醯CoA和肉毒鹼反應,生成輔酶A和脂醯肉毒鹼,脂肪醯基與肉毒鹼的3-羥基通過酯鍵相連接。催化此反應的酶為肉毒鹼脂醯轉移酶(carnitine acyl transferase)。線粒體內膜的內外兩側均有此酶,系同工酶,分別稱為肉毒鹼脂醯轉移酶I和肉毒鹼脂醯轉移酶Ⅱ。酶Ⅰ使胞漿的脂醯CoA轉化為輔酶A和脂肪醯肉毒鹼,後者進入線粒體內膜,同時酶Ⅰ將一分子基質中的肉鹼轉運至線粒體內膜外。位於線粒體內膜內側的酶Ⅱ又使脂肪醯肉毒鹼轉化成肉毒鹼和脂醯CoA,肉毒鹼重新發揮其載體功能,脂醯CoA則進入線粒體基質,成為脂肪酸β-氧化酶系的底物。
長鏈脂醯CoA進入線粒體的速度受到肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ的調節,酶Ⅰ受丙二醯CoA抑制,酶Ⅱ受胰島素抑制。丙二醯CoA是合成脂肪酸的原料,胰島素通過誘導乙醯CoA羧化酶的合成使丙二醯CoA濃度增加,進而抑制酶Ⅰ。可以看出胰島素對肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ有間接或直接抑制作用。飢餓或禁食時胰島素分泌減少,肉毒鹼脂醯轉移酶Ⅰ和酶Ⅱ活性增高,轉移的長鏈脂肪酸進入線粒體氧化供能。
β-氧化過程
脂醯CoA在線粒體基質中進入β氧化要經過四步反應,即脫氫、加水、再脫氫和硫解,生成一分子乙醯CoA和一個少兩個碳的新的脂醯CoA。
第二步加水(hydration)反應由烯醯CoA水合酶催化,生成具有L-構型的β-羥脂醯CoA。
第三步脫氫反應是在β-羥脂肪醯CoA脫飴酶(輔酶為NAD+)催化下,β-羥脂肪醯CoA脫氫生成β酮脂醯CoA。
第四步硫解(thiolysis)反應由β-酮硫解酶催化,β-酮酯醯CoA在α和β碳原子之間斷鏈,加上一分子輔酶A生成乙醯CoA和一個少兩個碳原子的脂醯CoA。
長鏈脂醯CoA經上面一次循環,碳鏈減少兩個碳原子,生成一分子乙醯CoA,多次重複上面的循環,就會逐步生成乙醯CoA。
從上述可以看出脂肪酸的β-氧化過程具有以下特點。首先要將脂肪酸活化生成脂醯CoA,這是一個耗能過程。中、短鏈脂肪酸不需載體可直拉進入線粒體,而長鏈脂醯CoA需要肉毒鹼轉運。β-氧化反應在線粒體內進行,因此沒有線粒體的紅細胞不能氧化脂肪酸供能。β-氧化過程中有FADH2和NADH+H+生成,這些氫要經呼吸鏈傳遞給氧生成水,需要氧參加,乙醯CoA的氧化也需要氧。因此,β-氧化是絕對需氧的過程。
脂肪酸β-氧化的生理意義 脂肪酸β-氧化是體內脂肪酸分解的主要途徑,脂肪酸氧化可以供應機體所需要的大量能量,以十六個碳原子的飽和脂肪酸硬脂酸為例,其β-氧化的總反應為:
CH3(CH2)14COSCoA+7NAD++7FAD+HSCoA+7H2O——→8CH3COSCoA+7FADH2+7NADH+7H+??
7分子FADH2提供7×1.5=10.5分子ATP,7分子NADH+H+提供7×2.5=17.5分子ATP,8分子乙醯CoA完全氧化提供8×10=80個分子ATP,因此一分子軟脂酸完全氧化生成CO2和H2O,共提供108分子ATP。軟脂酸的活化過程消耗2分子ATP,所以一分子軟脂酸完全氧化可凈生成106分子ATP。脂肪酸氧化時釋放出來的能量約有40%為機體利用合成高能化合物,其餘60%以熱的形式釋出,熱效率為40%,說明機體能很有效地利用脂肪酸氧化所提供的能量。
脂肪酸β-氧化也是脂肪酸的改造過程,機體所需要的脂肪酸鏈的長短不同,通過β-氧化可將長鏈脂肪酸改造成長度適宜的脂肪酸,供機體代謝所需。脂肪酸β-氧化過程中生成的乙醯CoA是一種十分重要的中間化合物,乙醯CoA除能進入三羧酸循環氧化供能外,還是許多重要化合物合成的原料,如酮體、膽固醇和類固醇化合物。
丙酸氧化
奇數碳原子脂肪酸,經過β-氧化除生成乙醯CoA外還生成一分子丙醯CoA,某些氨基酸如異亮氨酸、蛋氨酸和蘇氨酸的分解代謝過程中有丙醯CoA生成,膽汁酸生成過程中亦產生丙醯CoA。丙醯CoA經過羧化反應和分子內重排,可轉變生成琥珀醯CoA,可進一步氧化分解,也可經草醯乙酸異生成糖,反應過程見右圖。
α-氧化
脂肪酸在微粒體中由加單氧酶和脫羧酶催化生成α-羥脂肪酸或少一個碳原子的脂肪酸的過程稱為脂肪酸的α-氧化。長鏈脂肪酸由加單氧酶催化、由抗壞血酸或四氫葉酸作供氫體在O2和Fe2+參與下生成α-羥脂肪酸,這是腦苷脂和硫脂的重要成分,α-羥脂肪酸繼續氧化脫羧就生成奇數碳原子脂肪酸。α-氧化障礙者不能氧化植烷酸(phytanic acid,3,7,11,15-四甲基十六烷酸)。
ω-氧化
脂肪酸的ω-氧化是在肝微粒體中進行,由加單氧酶催化的。首先是脂肪酸的ω碳原子羥化生成ω-羧脂肪酸,再經ω醛脂肪酸生成α,ω-二羧酸,然後在α-端或ω-端活化,進入線粒體進入β-氧化,最後生成琥珀醯CoA。
不飽和脂肪酸氧化
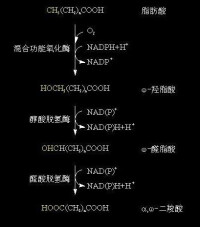
脂肪酸的w-氧化反應過程示意圖
不飽和脂肪酸完全氧化生成CO2和H2O時提供的ATP少於相同碳原子數的飽和脂肪酸。
生成的a,w-二羧酸可從兩端進行b-氧化作用而降解。動物體內的十二碳以下的脂肪酸常常通過w-氧化途徑進行降解;植物體內的在w-端具有含氧基團(羥基、醛基或羧基)的脂肪酸大多也是通過w-氧化作用生成的,這些脂肪酸常常是角質層或細胞壁的組成成分;一些需氧微生物能將烴或脂肪酸迅速降解成水溶性產物,這種降解過程首先要進行w-氧化作用,生成二羧基脂肪酸后再通過b-氧化作用降解,如海洋中的某些浮遊細菌可降解海面上的浮油,其氧化速率可高達0.5克/天/平方米。

基本信息
- 中文名
- 脂肪酸氧化
- 外文名
- fatty acid oxidation
- 特殊氧化方式
- 丙酸氧化、α-氧化、ω-氧化等
- 氧化方式
- β-氧化和特殊氧化方式
- 過程
- 活化、轉移、β氧化等
- 存在於
- 肝和肌肉,神經組織
- 來源
- 油脂水解