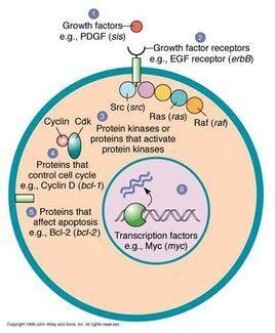
ras癌基因
ras癌基因
ras癌基因參與人類腫瘤的發生髮展,它最初是在急性轉化性逆轉錄病毒實驗中,從Harvey、Kirsten兩株大鼠肉瘤病毒中克隆出的轉化基因,自82年Weinberg等人發現其中有活化的H-ras基因后,引起了人們對ras癌基因在人類腫瘤發生、發展中所起作用的極大關注。經十餘年研究認為,ras癌基因參予細胞生長和分化的調控,參與多種腫瘤的形成與發展。
ras基因家族與人類腫瘤相關的特徵性基因有三種,即H-ras、K-ras和N-ras,它們分別定位於11、12、1號染色體,前二者是大鼠肉瘤病毒的轉化基因,後者是從人 神經母細胞瘤中分離得到的。
ras家族的基因所共有的特徵為:(1)基因組中均含有4個編碼的外顯子和1個5'端非編碼外顯子.(2)外顯子所編碼的蛋白為188-189個氨基酸殘基,分子量為21KD即P21 蛋白,此蛋白具有高度特異性和同源性,尤其在氨基酸序列的前80個氨基酸殘基中,幾乎無種屬間差別,具有高度保守性。
正常ras蛋白具有以下特點:(1)P21ras位於細胞膜的內側面,以軟脂酸共價鍵形 式固定於脂質雙層膜的內表面;(2)對GTP(三磷酸鳥苷)和GDP(二磷酸鳥苷)具有高度親 和力,並具有同源性GTP酶的活性.ras蛋白的這些生化性質與G蛋白非常類似,而G蛋 白具有從膜結合受體到腺苷酸環化過程的信號傳導作用,提示ras蛋白可能也具有信 號傳導通路的作用,但目前尚未發現ras蛋白作用的特異受體和靶細胞有人推測在哺 乳動物細胞中ras蛋白作用的靶分子很可能是磷脂酶C(磷脂酚肌醇,磷脂分解中的關 鍵酶)是生長信號傳遞途徑的一個重要成份,它被激活后可將4.5-二磷酸磷脂醯肌醇 (PIP2)裂解成三磷酸肌醇(IP3)和甘油二酯(DG),時IP3和DG是促進細胞增殖的第二信 使。現已證實,在ras癌基因轉化的細胞中,IP3,DG和PIP2的含量均明顯多於未轉化的細胞。
原癌基因的激活過程稱為活化,活化后的ras基因能使鼠NIH/3T3細胞系轉化為腫瘤細胞。在過去幾年中,對從腫瘤DNA中分離的活化ras基因,以及體外實驗中所獲得 的正常ras基因進行了較深入的研究,認為ras基因活化的主要方式有:(1)編碼區內的 突變.(2)插入激活。
ras基因家族的激活方式以點突變為主,在實體瘤中佔10~15%.較 公認的ras基因突變點位於三種ras基因的第12、13、59或61位氨基酸密碼子.Seeburg 等採用定點突變技術,對H-ras基因第12位密碼子一甘氨酸的所有置換氨基酸的可能性進行了分析,結果表明,此位點上,除置換氨基酸為脯氨酸外,其餘的置換氨基酸均具有轉化潛能,但有程度差異,到目前為止,在腫瘤中尚未發現二個或三個鹼基同 時突變。
在ras基因附近插入一個強起動子(Promoter)或增強子(enhaner)可使 ras基因表達增強。
正常基因的擴增或非編碼外顯子的缺失也能使ras基因呈高表達,但這兩種方式在ras基因激活中較少見。
4.活化ras蛋白的生化性質ras基因的任何突變使正常的ras蛋白轉變為能夠使 NIH/3T3細胞轉化的蛋白,從生化角度上講,這種突變激活可分為二種:(1)突變失去內在的GTP酶活性;(2)突變改變了對GDP和GTP的親和力.ras蛋白功能取決於編碼蛋白 的基因序列和與之密切相關的結構域。突變激活的常見位點多集中在GTP、GDP結合的結構域附近。這種結構的改變導致了ras蛋白的功能異常。目前認為:ras蛋白有活化和 非活化兩種形式,通常情況下,細胞內的ras蛋白分子處於非活化狀態。此狀態下的 ras蛋白特徵是具有能夠與GDP相結合的構象。在它們受到信號傳遞通道上游某一外界 因子的刺激時,便使GDP變成GTP,隨後ras蛋白髮生構象改變,成為其活化狀態。活化 的ras蛋白與效應分子相互作用,實現生長信號的傳遞。而且這種作用發生,活化的 ras蛋白會迅速失活,轉變為與GDP結合的非活化形式。此過程主要是ras蛋白自身的 GTP酶作用,它能催化GTP水解,使活化ras蛋白能立即轉變成為非活化狀態的ras蛋白.P21 蛋白通過與GDP和GTP結合併水解GTP完成其對正常細胞生長信號的傳遞和調節功能, ras基因的突變可擾亂正常狀態下活化與非活化ras蛋白的這種平衡機制,使正常非活 化的ras蛋白轉變成活化形式,實際上,ras基因的12、13、61位密碼子的活化突變常 不影響P21蛋白與GDP和GTP的結合,但卻降低了P21蛋白自身GTP酶活性,使其水解GTP 的速效大為降低,不斷激活靶分子,從而使ras蛋白維持於活化狀態,引起信號傳導 的持續效應,導致細胞大量增殖,而發生惡性轉化。從理論上講,使ras蛋白維持於活 化狀態有三種可能機制.(1)ras蛋白自身GTP酶活性降低,使GTP103水解減少;(2)GDP 與GTP間的交換增加;(3)某種機制誘導了不需與鳥嘌呤核苷酸結合的ras蛋白的活化構 象。還有人推測,ras蛋白的調節機制可能與其本身所具有的一種“開放”.“關閉”構型有關,構型處於“開放”時,能活躍地傳導特定的生長信號而“關閉”時則不能 傳導。如關鍵位點C12、13或61位密子)發生點突變,則使這種構型的開關不能再迴轉 到閉合狀態,使活化的ras蛋白持續傳導一種不適當的生長信號。
自82年以來,已經在膀胱癌、乳腺症、結腸癌、腎癌、肝癌、肺癌、胰腺癌、胃癌、及造血系統腫瘤中,均檢測出了ras癌基因的異常.Pulciani等的研究結果表明,被檢腫瘤DNA中所含活化ras基因僅佔10~20%,似乎ras癌基因在人類腫瘤發生髮展中 並非起主要作用。事實上,ras癌基因參予多種腫瘤的發生髮展,只不過突變率相差很 大.(1)不同腫瘤類型,ras癌基因的突變率相差明顯,如最高可達90%(胰腺癌),其次 是甲狀腺癌(53%)和結腸癌(47%).(2)突變ras癌基因的種類與某些腫病類型密切相 關,即有優勢激活現象。如胰腺癌、結腸癌、肺癌等以K-ras突變為主,造血系統腫瘤 多發現N-ras的突變,泌尿系腫瘤則以H-ras突變為主,目前的資料表明,rasH和rasK 的表達不僅與膀胱癌、腎盂癌、肺癌、結腸癌、有密切的關係,而且也與膽囊癌、胰 腺癌、腎母細胞癌、慢性淋巴細胞白血病,黑色素瘤形成密切相關,N-ras表達水平 上升雖然主要發生在造血系統的惡性腫瘤中,如APL、AML、BL,但在神經母細胞瘤、纖維肉瘤,橫紋肌肉瘤中的表達也有一定的上升,並認為是這些腫瘤形成的主要原 因。因此,檢測ras突變對了解腫瘤的發生髮展,以及監測惡性腫瘤的治療效果具有 重大意義,對臨床工作具有重要指導意義。