谷氨酸受體
谷氨酸受體
谷氨酸受體分為兩類:一類為離子型受體,包括:N-甲基-D-天冬氨酸受體(NMDAR)、海人藻酸受體(KAR)和α-氨基-3 羥基-5 甲基-4 異惡唑受體(AMPAR),它們與離子通道偶聯,形成受體通道複合物,介導快信號傳遞;另一類屬於代謝型受體(mGluRs),它與膜內G-蛋白偶聯,這些受體被激活后通過G-蛋白效應酶、腦內第二信使等組成的信號轉導系統起作用,產生較緩慢的生理反應。
離子型受體
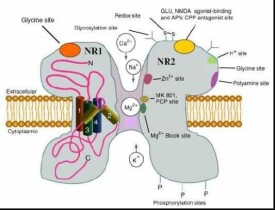
(1) NMDA 受體(NRs):其與突觸的可塑性和學習記憶密切相關。通過該受體本身、其共軛的離子通道及調節部位3 者形成的複合體而發揮功能,對Ca2+高度通透。每個NMDA 受體上含有兩個谷氨酸和兩個甘氨酸結合識別位點,谷氨酸和甘氨酸均是受體的特異性激活劑。到目前為止已克隆出5個亞基,NMDAR1、NMDAR2(A-D)其中NMDAR1 可單獨形成功能性純寡聚體NMDAR,但NMDAR2 亞基卻不具備該功能。有研究表明NMDAR可能是由NMDAR1 和NMDAR2 不同的亞基組成的一個異寡聚體。
(2) KA/AMPA 受體:它們也是受配基調控的離子通道,對Na+、K+有通透性,研究證明,一些受體亞型對Ca2+也有通透性。AMPA 家族包括4 個結構極為相似的亞基GLUR1-4,各亞基的氨基酸序列的同源性高達70%。由於氨基酸殘基的疏水性分佈,在靠近羧基端的部分構成4 個跨膜區。AMPA、L-谷氨酸及KA 均可激活這類離子通道,並有AMPA 的高親和力結合位點。天然的AMPAR 是由這4 種亞基形成的四聚體。每個單位的分子量為108kd。AMPA 受體的4 種亞基在第4 個跨膜區上游均含有1 個由38 個氨基酸殘基組成的特殊區段,該區存在2 個結構相似區,分別由受體基因上的2 個相臨的外顯子編碼。但各亞基的DNA 編碼在翻譯后要經過一些如:磷酸化、糖基化等修飾,這些修飾是通道功能的重要調節方式。離子型谷氨酸受體功能的多樣性是通過不同亞基組裝、選擇性基因結合和轉錄前mRNA 的編輯等方式來實現的。
在大鼠中通過分子克隆技術,已發現5 種KA 受體亞型(GLUR5-7、KA-1、KA-2),利用逆轉錄PCR 及膜片鉗技術揭示:KA 受體是由同類的不同亞基組成的異質組合體。亞基的組成對受體的功能和特性影響特別大,因為異質的KA 複合物中出現編輯的GLUR5 或GLUR6 會阻礙Ca2+的通透性。細胞可能通過 RNA 編輯改變結構,達到調控通道的Ca2+流量。
代謝型谷氨酸受體(mGLuRs)
這是通過G-蛋白偶聯,調節細胞內第二信使的產生而導致代謝改變的谷氨酸受體,其可分為不同的8 個亞型mGLUR1-8,根據氨基酸序列的同源性及其藥理學特徵和信號轉導機制的不同,可將其分為3 組,ⅠmGLUR1、mGLUR5; Ⅱ mGLUR2-3; Ⅲ mGLUR4、mGLUR6-8。Ⅰ組可被Quis 強烈活化並與磷脂酶C 途徑(PLC)相偶聯;Ⅱ、Ⅲ組均可與腺苷酸環化酶系統(AC)被動偶聯。
D-Ser-NMDA受體的一種新的調控因子 陳福俊
陳福俊,何德富,周紹慈
(華東師範大學上海市腦功能基因組學重點實驗室,上海 200062)
摘要:最近研究證實哺乳動物神經系統中存在內源性的D-Ser。這種內源性D-Ser在神經系統中的分佈與NMDA受體的分佈相平行,進一步的研究表明,D-Ser由突觸旁星形膠質細胞產生,而作用於突觸后NMDA受體上的Gly結合位點,對NMDA受體的功能進行調控。本文將綜述D-Ser在神經系統中的分佈、合成及其生理機能。
Abstract: The viewpoint that there is no endogenous D-serine in mammalian nervous system has changed based on recent published reports. The study indicated that the distribution of this kind of endogenous D-serine parallels with that of NMDA receptor in nervous system. Further study suggested that the D-serine produced in astrocytes regulates the function of NMDA receptor through the glycine-binding site of the receptor. This paper reviews the distribution, synthesis and physiological function of D-serine in nervous system.
NMDA受體(N-methyl-D-aspartate receptor)是中樞神經系統內一類重要的興奮性氨基酸(excitatory aminoacid, EAA)受體。NMDA受體不僅在神經系統發育過程中發揮著重要的生理作用,如可調節神經元的存活,調節神經元樹突、軸突結構發育及參與突觸可塑性的形成等;在神經元迴路的形成中NMDA受體亦起著關鍵作用,有資料表明NMDA受體是學習與記憶過程中一類至關重要的受體。NMDA受體受多種因子的調控,而最近發現了NMDA受體的一種特別的調控因子——D型絲氨酸(D-Serine,D-Ser)。D-Ser這種新的神經調質的發現引起了神經科學界極大關注,本文將綜述這方面的最新進展。
自然界中存在的氨基酸絕大多數都有兩種構型:L型和D型。構成生物體中蛋白質的氨基酸通常都為L型,在細菌和無脊椎動物體內曾發現有內源性的D型氨基酸,而對於哺乳動物體內存在D型氨基酸的事實,過去普遍把它歸結於食物來源或腸內的細菌所產生[3,4]。早期研究認為,哺乳動物體內不能產生D型氨基酸,因為氨基酸消旋酶只發現在細菌和昆蟲體內,而在哺乳動物體內還未分離純化出這種酶。然而最近的研究報告表明在哺乳動物神經系統中存在自由D-Ser和D-天冬氨酸(D-aspartate, D-Asp)[5,6];另有研究報告報道,大鼠額葉和頂葉皮層突觸體中的磷酸絲氨酸磷酸化酶(phosphoserine phosphatase)可水解L-磷酸絲氨酸(L-phosphoserine),最終生成L-Ser和D-Ser。因而這類D型氨基酸在神經系統中的生理作用,很自然地引起了神經科學工作者的廣泛關注。下面將重點闡述D-Ser在神經系統中的分佈、合成及生理機能。
自有報道說在哺乳動物中樞神經系統中存在自由的D型氨基酸以來,許多科學家便把注意力轉向這類不參與構成生物體內蛋白質的氨基酸上來。採用對D-Ser具有高度選擇性的抗體,利用免疫組化的方法研究發現,D-Ser主要存在於哺乳動物腦內灰質區中的Ⅱ型星形膠質細胞內,這種膠質細胞多位於突觸旁邊[2,8]。D-Ser分佈的主要腦區為前額葉皮層和紋狀體,小腦中也有微量分佈。令人感興趣的是,在哺乳動物中,NMDA受體分佈密度最高的是海馬的CA1區、CA3區和齒狀回,大腦皮層區的前腦皮層、前扣帶區和梨狀皮層,此外,在紋狀體、丘腦、小腦顆粒細胞層也有較多分佈。在D-Ser存在的腦區中,往往存在著大量的NMDA受體,二者的分佈存在著一種平行關係。
NMDA受體是一種獨特的雙重門控通道(doubly gated channel),它既受膜電位控制也受其它神經遞質控制。NMDA受體被激活后,主要對Ca2+有通透性,介導持續、緩慢的去極化過程。在突觸傳遞過程中,NMDA受體的激活需要非NMDA受體的參與,其中主要是AMPA受體(α-amino-3-hydroxy-5-methyl-4-isoxazole propionate receptor)的參與。當刺激達到一定強度時,突觸前膜釋放的谷氨酸作用於AMPA受體,通過AMPA受體通道的離子流增強,使得鄰近NMDA受體的突觸后膜局部去極化,進而導致NMDA受體通道Mg2+阻斷的釋放,這時谷氨酸與NMDA受體的結合便可使通道打開。此外當有甘氨酸結合到甘氨酸結合位點時,通過變構調控可以大大增強谷氨酸作用於NMDA受體后所產生的效應,另外多聚胺可增強谷氨酸對NMDA受體的作用,而Zn2+卻可以抑制多聚胺的這種作用。可見,NMDA受體的激活受多種因子的調控。
D-Ser與NMDA受體的共分佈現象,促使科學家進一步對二者的功能聯繫進行研究。最近的研究發現D-Ser是NMDA受體的一種新的獨特的調質。Ascher等人研究發現,在用快速灌流製備的神經標本中,NMDA受體的活性會暫時喪失,但這種活性的暫時喪失可為甘氨酸所逆轉。進一步的研究表明NMDA受體上存在Gly結合位點,而NMDA受體的活化需要Gly位點的共激活。但讓人不解的是,在中樞神經系統中Gly的濃度在前腦是最低的,而前腦中NMDA受體的含量卻很高;相反在脊髓和後腦中Gly的含量最高,但在這些區域Gly卻發揮著一種抑制性神經遞質的作用。有研究報告顯示在NMDA受體分佈的灰質腦區中,D-Ser的含量約為Gly的3倍[12-14],在前額葉皮層,胞外D-Ser的量與Gly相當,在紋狀體D-Ser的含量約是Gly的2倍。另外的研究證實,施加外源性的D-Ser可以增強NMDA受體調節的神經傳遞作用。此外採用放射性同位素3H標記D-Ser研究發現,D-Ser選擇性結合於Gly結合位點。大量實驗結果表明,D-Ser對NMDA受體似乎與Gly有著類似的作用,D-Ser也是NMDA受體上Gly位點的一種內源性配基。進一步的研究則驚人的發現D-Ser是由突觸旁星型膠質細胞產生而作用於突觸后神經元上NMDA受體的。如Fig.
1所示,當突觸前膜興奮時,釋放興奮性氨基酸遞質谷氨酸到突觸間隙,其中一部分Glu作用於突觸后膜上的NMDA受體,另一部分則擴散到了鄰近的星形膠質細胞,作用於其上的非NMDA受體,後者的激活導致貯存於胞漿中的D-Ser由星形膠質細胞釋放出,然後結合於突觸后膜上的NMDA受體,從而與Glu共同作用,激活NMDA受體。
Fig. 1 The active mechanism of D-serine(cited from Snyder SH, revised partly)
而D-Ser的清除和滅活則依賴於D型氨基酸氧化酶,在三十年代便發現哺乳動物體內存在D型氨基酸氧化酶,但對其作用一直沒有定論。D型氨基酸氧化酶是一種高度選擇性的酶,只降解中性D型氨基酸。Snyder等採用組織化學的方法研究發現,D型氨基酸氧化酶在腦內的分佈存在區域差異,其濃度與D-Ser的分佈相平行。在生理條件下D型氨基酸氧化酶的最佳底物是D-Ser和D-丙氨酸,在腦片實驗中也證實了D型氨基酸氧化酶可降低NMDA受體的神經傳遞作用,而對Gly和D-Asp無明顯影響,表現出對D-Ser的高度選擇性。可見通過D型氨基酸氧化酶的作用可及時清除釋放入突觸間隙的多餘的D-Ser,以防止其持續作用於NMDA受體,從而維持了NMDA受體的正常生理功能。
D-氨基酸的產生依賴氨基酸消旋酶,胞內是否存在專一的絲氨酸消旋酶呢?Snyder等的工作使這一問題獲得了圓滿的解答,他們從大鼠腦內分離出一種氨基酸消旋酶,這種消旋酶專一催化L-Ser轉化為D-Ser,遂將其命名為絲氨酸消旋酶(serine rasemase)。絲氨酸消旋酶由339個氨基酸殘基組成,分子量為36.3KD。爾後他們通過分析絲氨酸消旋酶的結構和檢索基因庫后,得出這一酶的全序列cDNA,並在體外進行了表達,而且成功地在培養細胞中合成了D-Ser。免疫組化研究表明,這種絲氨酸消旋酶大量存在於前腦及灰質區的Ⅱ型星形膠質細胞中,而這裡正是NMDA受體和D-Ser分佈廣泛的腦區。這也進一步說明D-Ser是生物體內源性的物質。
以往被認同的神經遞質和神經調質都是由突觸前膜或突觸后膜產生,作用於突觸前膜或突觸后膜上的受體,而D-Ser這種獨特的神經調質則是由突觸旁的神經膠質細胞產生作用於突觸后膜上的NMDA受體,這就使我們想到腦內數量遠多於神經元的神經膠質細胞,除了能高親和性地吸收GABA並將其滅活,通過細胞間液的物質交流參與神經元內Glu和GABA的代謝,影響神經肽的合成及傳統的支持、營養、修復、吞噬諸多功能外,還可能參與神經興奮性傳導和調製等機能。
基本信息
- 中文名
- 谷氨酸受體
- 種類
- 離子型受體