細菌域
細菌域
Woese依據16S rRNA序列上的差別,認為這兩類生物和真核生物一起從一個具有原始遺傳機制的共同祖先分別進化而來,因此將三者各置為一“域”(Domain),作為比界高的分類系統,並分別命名為細菌域(Bacteria)、古菌域(Archaea)和真核域(Eukarya)。
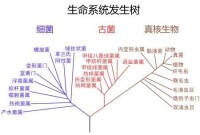
生命系統發生樹
細菌廣泛分佈於土壤和水中,或著與其他生物共生。人體身上也帶有相當多的細菌。據估計,人體內及表皮上的細菌細胞總數約是人體細胞總數的十倍。此外,也有部分種類分佈在極端的環境中,例如溫泉,甚至是放射性廢棄物中,它們被歸類為嗜極生物,其中最著名的種類之一是海棲熱袍菌(Thermotoga maritima),科學家是在義大利的一座海底火山中發現這種細菌的。然而,細菌的種類是如此之多,科學家研究過並命名的種類只佔其中的小部份。細菌域下所有門中,只有約一半能在實驗室培養的種類。
細菌的營養方式有自營及異營,其中異營的腐生細菌是生態系中重要的分解者,使碳循環能順利進行。部分細菌會進行固氮作用,使氮元素得以轉換為生物能利用的形式。細菌也對人類活動有很大的影響。一方面,細菌是許多疾病的病原體,包括肺結核、淋病、炭疽病、梅毒、鼠疫、砂眼等疾病都是由細菌所引發。然而,人類也時常利用細菌,例如乳酪及酸奶的製作、部分抗生素的製造、廢水的處理等,都與細菌有關。在生物科技領域中,細菌有也著廣泛的運用。
古細菌域
古細菌原本是被認為是一些可生活於極嚴酷環境的細菌。如在溫泉水中,濃鹽水中,酸、鹼性極高的水中,或是礦井的水中等等。生物學家對於它們的適應性感到驚奇,以為它們僅是少數的情況。
但後來發現它們的存在十分廣泛,甚至在海水中亦分佈甚廣,於是正視它們的存在,並認同為可獨立成一類的生物。但由於研究不足,生物學家們對它們的認識不足,所以它們的分類尚未成熟。現只能確認它們當中的四個門。其它的還有甚多尚待研究及確定。
最早發現的古核生物是產甲烷細菌類。但隨著進化分子生物學與進化細胞生物學的興起,特別是發現在海洋深處的熱泉口很高溫度的環境中存在眾多嗜熱細菌,是人們很自然聯想到這些古細菌與地球早期生命環境的關係,設想它們可能在細胞起源與進化中扮演過重要角色。
古核生物細胞的形態和遺傳結構裝置與原核細胞相似,但有些分子進化特徵更接近真核細胞,有些生物學家建議將生物分為原核生物、古核生物與真核生物3大領域,將其細胞相應分為3大類型:原核細胞、古核細胞和真核細胞。
古細菌體積小,直徑約 0.5-5μm, 形狀有桿狀、球形、螺旋形、絲狀,基因組大小為 2-4mbp,多數為嗜熱、嗜酸、自養型,古細菌有豐富的組蛋白,DNA 以核小體的形式存在。古細菌與一般細菌的氧化磷酸化產能不同, 如產甲烷菌(Methanogen)和硫細菌(Sulfolobus)用氫還原 CO2、S生成 CH4、H2S 來獲得能量, 有些古核生物利用鎢代替高溫下不穩定的 NADH 參與電子傳遞鏈。古細菌具有比細菌更多樣化的質膜結構。古細菌質膜由特殊脂類組成, 甘油和脂肪酸鏈之間的結合是通過醚鍵而不是酯鍵來實現的。古細菌沒有以胞壁酸為基質的粘肽類細胞壁,而真細菌具有與胞壁酸相結合的細胞壁。古細菌對作用於真細菌細胞壁的抗生素如青霉素、頭孢黴素以及抑制真細菌翻譯過程的氯黴素和環絲氨酸不敏感, 而對抑制真核生物翻譯過程的白喉毒素卻十分敏感。與真細菌相比, 不同類型古細菌的多種蛋白質一級結構中疏水殘基的平均數量增多, 增強了蛋白質的高溫穩定性。古菌、細菌及真核生物在細胞結構和基因組結構方面的異同。
古細菌域 - 古細菌與真核細胞遺傳信息傳遞機制相似 古細菌比真細菌更可能真核細胞的祖先,二者曾在進化上有過共同歷程。
1.細胞壁的成分 真細菌的細胞壁主要由含壁酸的肽聚糖構成,古細菌也有細胞壁,但其成分卻與真細胞一樣完全不含上述成分,因此抑制壁酸合成的鏈黴素,抑制肽聚糖前體合成的環絲氨酸,抑制肽聚糖合成的青霉素與萬古黴素等對真細菌類有強的抑制生長作用的藥物對古細菌與真核細胞無作用。
2.DNA與基因結構 真核細胞DNA中含有大量的重複序列,真細菌的細胞DNA不含重複序列,而古核細胞DNA中卻有重複序列存在。
3.核小體結構 原核細胞核區主要由一個環狀DNA分子組成,沒有或只有極少的蛋白質與DNA結合,古核細胞具有組蛋白,而且能與DNA構建成類似核小體的結構,但與真核細胞典型的核小體又有差異。
4.核糖體多數古細菌類的核糖體介於真核細胞與真細菌之間。古細菌對作用於真細菌核糖體大、小亞基,抑制蛋白的抗生素不敏感, 而對抑制真核生物翻譯過程的白喉毒素卻十分敏感。
5.5S rRNA根據對 5S rRNA 序列分析及二級結構的研究表明古細菌與真核生物相似,與原核生物差距甚遠。
除上述各點外,根據DNA聚合酶分析,氨醯- tRNA 合成酶的作用,起始氨醯tRNA與肽鏈延長因子等分析,也說明古細菌與真核生物在進化上的關係較原核生物更為密切。
古細菌域包括一些嗜熱和超嗜熱古細菌、極端嗜鹽古細菌、產甲烷古細菌和熱原體古細菌,分為兩個門,共有217個種。目前,可在實驗室培養的古菌主要包括三大類:產甲烷菌、極端嗜熱菌和極端嗜鹽菌。