分子進化
生物進化過程生物大分子的演變現象
分子進化
molecular evolution
生物進化過程中生物大分子的演變,包括前生命物質的演變;蛋白質分子和核酸分子的演變以及細胞器和遺傳機構(例如遺傳密碼)的演變。分子進化的研究可以為生物進化過程提供佐證,為深入研究進化機制提供重要依據。
廣義的分子進化有兩層含義,一是原始生命出現之前的進化,即生命起源的化學演化;二是原始生命產生之後生物在進化發展過程中,生物大分子結構和功能的變化以及這些變化與生物進化的關係,這就是通常所說的分子進化。
地球的歷史約有50億年。60年代末在南部非洲的前寒武紀地層中發現了一種古老的細菌化石,它的生存年代約在32億年之前,另外還發現一些更古老的類似原藻類的微小生物化石,它的生存年代約在34億年以前,這是地球上發現得最早的生命的記錄。
關於地球以及其他天體上的生命進化的研究方面,1936年蘇聯生物化學家 A.И.奧帕林提出的團聚體理論、1952年英國生物物理學家J.D.貝爾納提出的粘土表面理論、1959年美國學者S.W.福克斯和K.原田馨提出的類蛋白微球體理論、1969年江上石二夫提出的海生顆粒理論等都各自具有獨特的見解。1975年美國物理化學家和生物化學家M.卡爾文在總結各家學說的基礎上提出一個模型,認為最初覆蓋於地球上的那些致生元素先是形成各種原始的致生分子(甲烷、硫化氫等)。致生分子在許多種能源(包括太陽的紫外線、電離輻射能和隕石衝擊波等)的影響下進一步形成低分子有機化合物,以後再從低分子有機化合物過渡到高分子有機化合物。大約在40億年前才由大分子有機化合物形成了最初的具有生命形態的有機體。
關於生命起源的原始環境條件,奧帕林曾有專著論述。第一個嘗試用實驗論證奧帕林理論而獲成功的是美國人H.C.尤里和S.L.米勒。他們在1952年首先模仿生命前時期的環境條件,在甲烷、氨、氫和水的混合物中通過放電反應形成多種產物,包括有各種氨基酸、嘌呤、嘧啶和一些簡單的糖類分子。以後在另一些條件下發現核苷的磷酸化現象。從1968年以來人們發現在星際空間同樣存在有類似的有機化合物分子,在太空隕石以及月球塵埃中有些跡象顯示甚至有氨基酸的存在。1958年福克斯的實驗證實了無水氨基酸混合物在高於 100℃的溫度下縮合成為類蛋白。類蛋白在水和高濃度的鹽溶液中能形成直徑約0.5~3微米的微球體。微球體甚至能以出芽的方式進行“繁殖”。核酸同樣也可以在模擬的實驗室條件下由核苷酸形成。所有這些實驗結果都說明生物大分子可以在原始的地球表面不通過酶促反應而在生物體外形成。蛋白質和核酸在前生命進化階段中哪一種出現在先以及他們的相互依賴關係究竟怎樣,都是研究和爭論中的問題。
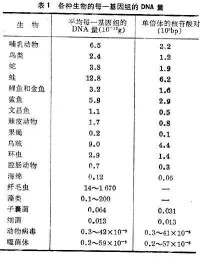
核酸是遺傳物質,可以明顯地看到在生物進化過程中各種生物每一基因組的核酸的量在總的趨勢上逐漸增加(表1)。
從總的趨勢來看,愈是低等生物DNA量愈少,愈是高等則愈多。但是這規律對於某些生物顯然並不適用,原因是多方面的。一般生物愈是高等則所需要的基因愈多(見基因),可是進化達到某一階段以後,基因的數目便不再相應地增加。細菌的呼吸代謝和氨基酸、核苷酸代謝途徑與人沒有多大差別,這一事實足以說明有關的酶的結構基因沒有太大的增加。倍性對於每一細胞中的DNA的含量有很大的影響。纖毛蟲的DNA含量特別高便是由於這一原因。例如雙小核草履蟲(Paramecium aurelia)的大核是860倍體,梨形四膜蟲(Tetrahymena pyriformis)的大核是100倍體。被子植物的每一細胞中的DNA含量較高也是由於這一原因(見染色體倍性)。各種生物的不編碼蛋白質的重複序列和內含子的量不同是使 DNA含量不同的另一原因。
生物進化過程中 DNA的質也在發生變化。用分子雜交方法可以分析各種生物的DNA的相似程度(表2)。
對於某一類生物來講,例如在靈長類動物和細菌等生物中都可以用同樣的方法來測定它們的親緣關係(圖1)。
進化中的保守性 分子雜交測定的結果只能說明兩種生物的DNA的相同或不同程度,通過DNA順序分析才能知道它們怎樣相同或不同。對於大腸桿菌和λ噬菌體等的46個啟動區進行核苷酸順序分析,發現在每一個基因的mRNA轉錄位置前面相隔10個鹼基對的地方有一個稱為普裡布諾順序的保守區,它包括核苷酸順序TATAATG。這裡面特別是從左面開始第1、第 2和第6這幾個鹼基如果變為G或C就會導致轉錄效率下降,尤其是第6位上的T,在46個例子中從沒有發現過例外。在高等生物中存在著的類似保守區稱為霍格內斯順序。
在 5個腸道桿菌科(Enterobacteriaceae)細菌中曾經分析了它們的染色體DNA複製起點的245個核苷酸的順序,發現它們的相同的順序可以多達85%,同樣說明這一順序具有很強的保守性。
這一類保守區的存在說明這些結構對於各自的功能是十分重要的,因而在生物進化過程中不容輕易改變。至於個別基因或染色體片段的位置改變則在進化過程中可以發生而保存下來。個別核苷酸的改變同樣可以發生而保存下來,這些變化可以清楚地反映在各種近緣生物的染色體和遺傳學圖的比較研究中,也可以反映在蛋白質的比較研究中。
蛋白質差異 可以用免疫學方法測定各種生物的蛋白質的親緣關係,例如用人的清蛋白注射家兔,從家兔取得抗血清,把抗血清分別和人、大猩猩、黑猩猩等的清蛋白進行沉澱反應測定,可以看到愈是親緣關係相近的清蛋白沉澱反應愈強。
同工酶的電泳測定是70年代發展起來的可以用來比較生物蛋白質的親緣關係的方法。同工酶是功能相同而一級結構不相同的酶。一種蛋白質中任何一個氨基酸的替代,只要它帶來用電泳方法可以區分的電荷差別就可被檢出,但如果沒有帶來電荷差別便不能檢出。由於這一方法簡便、快速,所以在分子進化的研究中常被採用。例如曾用電泳方法對包括魏氏果蠅 (Drosophila wil-listoni)在內的9種果蠅14個亞種的 36種酶的同工酶進行分析,根據分析的結果可以繪製出它們的系譜圖。
氨基酸分析結果可以為生物的親緣關係的研究提供更多的資料。細胞色素 C是從人到酵母菌中都存在的一種蛋白質,便於進行廣泛的比較。細胞色素C由140個氨基酸構成。把各種生物的細胞色素 C的氨基酸成分和人的相比較,可以看到親緣關係愈近的生物的細胞色素 C和人的愈相近似(表3)。在血紅蛋白、血纖維蛋白中都進行過同樣的研究。根據多種動物中這三種蛋白質的氨基酸成分的研究結果,還可以看到生物進化過程中細胞色素C的進化速率最低。纖維蛋白的進化速率最高,血紅蛋白則介於兩者之間(圖2),也就是說細胞色素 C是最保守的蛋白質。例如人和獼猴的共同祖先生活在4000~5000萬年以前,而到現在這樣長的時間內,細胞色素C中只有一個氨基酸發生了變化(表3)。染色體中的組蛋白是一類更為保守的蛋白質。
成人的血紅蛋白由兩個 α鏈和兩個β鏈構成,胎兒的血紅蛋白由兩個 α鏈和兩個γ鏈構成。α、β和γ鏈都由146個氨基酸構成,而且許多氨基酸都是相同的,並且和肌紅蛋白也非常近似。根據對於許多種動物的血紅蛋白和肌紅蛋白的研究,可以畫出血紅蛋白分子的進化樹(圖3)。
在漫長的進化過程中生物的 DNA經歷了各種各樣的變化。包括基因突變、基因重組、染色體易位等。
鹼基置換突變常導致蛋白質中一個氨基酸的改變。例如正常血紅蛋白第 6位的谷氨酸改變為纈氨酸便成為鐮形細胞貧血症的血紅蛋白 HbS,為賴氨酸替代則成為HbC,前者的鹼基是從GAA(谷氨酸)→GUA(纈氨酸),後者是 GAA(谷氨酸)→AAA(纈氨酸)。已經發現的由α鏈上單個氨基酸的改變而造成的異常血紅蛋白不下40種,由β鏈上單個氨基酸改變而造成的異常血紅蛋白在80種以上。
DNA 分子的重複也是蛋白質分子發生改變的重要原因。例如血紅蛋白中的 α、β、γ、δ 4種多肽鏈相互關係便是如此(圖3)。
缺失在蛋白質分子進化中經常發生。例如血紅蛋白第22位殘基根據不同類型多肽鏈的前後順序對應比較,可以肯定人和馬的α鏈的差別由一個缺失造成。
還有一種情況是鏈的延伸,例如血紅蛋白HbCS是一種具有正常 α鏈的變種,它從正常 α鏈羧基端的精氨酸上又延伸出31個附加的氨基酸殘基,成為具有172個氨基酸的肽鏈,附加的氨基酸序列與 α鏈的其他部分或其他的血紅蛋白鏈毫無關係。
遺傳密碼的進化 現有的生物的核基因密碼都是相同的(見遺傳密碼),說明遺傳密碼體系在生物進化過程中早已固定。
1979年在三個實驗室中分別發現遺傳密碼的統一性原則不適用於線粒體基因。例如人和酵母菌的線粒體中的UGA並不是終止密碼子而是色氨酸密碼子,AUA在人的線粒體中不是異亮氨酸而是甲硫氨酸密碼子。這些事實說明在生物進化過程中遺傳密碼確實發生過變化。
哺乳動物的線粒體DNA的全部順序已經分析清楚,這裡面可以找到有23個編碼轉運核糖核酸(tRNA)的順序。酵母菌的線粒體 DNA用順序分析和分子雜交方法也得到和哺乳動物線粒體大約相同數目的tRNA順序。染色體基因所編碼的tRNA至少有50種。線粒體中的tRNA種類少於染色體基因所編碼的tRNA種類。這一事實說明線粒體的密碼體系是比較原始的狀態。
對於各種生物物種的每一個蛋白質,用每一個位點每年發生的氨基酸替換的次數為標準衡量分子進化的速率是大致恆定的,只要該分子的功能和三維結構保持不變。
功能上較次要的分子或分子的區域的進化速率(按突變替換數/每位點/每年計算)要比功能重要的分子或分子的部分的進化快。
對現存分子的結構或功能破壞較小的那些突變替換(即保守性替換)要比破壞力較大的突變替換的進化來得頻繁。
基因複製總是在獲得一個新功能之前就已發生。

基本信息
- 中文名
- 分子進化
- 外文名
- molecular evolution
- 簡介
- 生物進化過程中生物大分子的演變
- 前生命進化
- 地球的歷史約有50億年