暴龍超科
暴龍超科
暴龍超科(Tyrannosauroidea)是肉食性恐龍的一個超科或演化支,屬於獸腳亞目的虛骨龍類,並包含暴龍科與其更原始的近親。暴龍超科最初出現於侏羅紀的勞亞大陸。到了白堊紀,它們已經成為北半球的大型優勢食肉動物,暴龍則是當中最大型的物種。暴龍超科的化石已發現於西歐、北美洲、中亞、以及東亞等地,可能還有澳洲。
如同大部分的獸腳類恐龍,暴龍超科是群二足、肉食性恐龍,並具有許多的共有衍征,尤其是頭顱骨與骨盆。在暴龍超科剛出現時,它們是群小型掠食動物,前肢細長,有三根手指。晚白堊紀物種的體型更為龐大,有數種則成為有史以來陸地上最大型的掠食動物之一;但大部分後期物種的前肢很短小,並具有兩指。帝龍是種早期的暴龍超科動物,出土於中國,已發現具有原始羽毛,其他暴龍超科可能也具有羽毛。許多暴龍超科的頭部具有骨質冠,形狀與尺寸變化很大,可能具有視覺辨識功能。
暴龍超科這名詞的使用,主要用在提及暴龍類的原始物種時。

霸王龍
早期暴龍超科動物的頭顱骨較長、低矮、輕型,類似其他的虛骨龍類,而晚期物種的頭顱骨較高、大型。儘管頭顱骨有者外型上的差異,但所有暴龍超科具有特定的頭部特徵。前上頜骨非常高,口鼻部前端呈鈍狀,這是與阿貝力龍科平行演化的特徵。鼻骨互相固定者,並往上稍微拱起,形成頭部上側表面的隆起。前上頜骨的牙齒小於與其他部位的牙齒、形狀也不同、橫剖面為D形。除了原始的冠龍,暴龍超科下顎的上隅骨有個明顯的棱脊,從顎部關節下方往兩旁隆起。
如同大部分其他的獸腳類恐龍,暴龍超科具有S形彎曲的頸部、長尾巴。早期的物種具有長前肢,冠龍的前肢長度約是後肢的60%;早期物種具有3根手指,是典型的虛骨龍類特徵。早白堊紀的始暴龍仍保有長前肢。但不清楚阿巴拉契亞龍是否也具有長前肢。更衍化的暴龍科,前肢大幅地縮小,最極端的例子為蒙古的特暴龍,它們的肱骨只有股骨的1/4長度。前肢的第三指也隨者時間而逐漸縮小。原始的冠龍的第三指並未縮小,但帝龍的第三指已經明顯地較其他兩指細。始暴龍仍保有三根具有功能的手指,而暴龍科動物只有兩指,不過已在某些標本上發現了退化的第三指遺跡。如同大部分的虛骨龍類,它們的第二指是最長的一根,無論第三指是否存在。
暴龍超科的骨盆特徵包含:腸骨尾端前上側有個凹處、腸骨外側有個銳利的垂直棱脊,從臀窩處往下延伸、恥骨尾端巨大,佔了恥骨把柄部的過半長度。所有的暴龍超科都具有這些特徵,包含原始的冠龍與帝龍。祖母暴龍與史托龍的恥骨沒有被發現,但兩者的腸骨都符合暴龍超科的特徵。
如同大部分獸腳類恐龍,所有暴龍超科的後肢具有四個腳趾,而第一趾沒有接觸地面。暴龍超科的後肢與身體比例,大於幾乎所有其他的獸腳類恐龍;它們的後肢具有一些善奔動物的特徵,包含修長的脛骨與蹠骨。最大型的成年暴龍依然保有類似的腿部比例,但暴龍推斷無法奔跑。暴龍科的第二、第四蹠骨夾住第三蹠骨,形成夾蹠型態。阿巴拉契亞龍也具有夾蹠型態腳踝,但始暴龍則不確定是否也有夾蹠型態腳踝。似鳥龍科、傷齒龍科、近頜龍科也具有夾蹠型態腳踝,而暴龍超科的原始成員則無此特徵,例如帝龍,顯示這為平行演化的結果。
--Tyrannosauroidea暴龍超科=={暴龍>似鳥龍,今鳥亞綱}=Tyrannosauria=Deinodontoidea恐齒龍超科
| 未定:
| 未命名種 (上頜骨 牙)--早 Cretaceous--日本
| ?Futabasaurus千葉龍
|--Eotyrannus始暴龍
|?-Iliosuchus髂鱷龍

暴龍超科
|?-Stokesosaurus史托龍
|?-Tonouchisaurus吐魯茨龍
`--Tyrannosauridae 暴龍科=Deinodontidae恐齒龍科
| 未定:
| 未確定化石 材料:中-晚Campanian--加拿大艾伯塔
| Chingkankousaurus金剛口龍
|--Alectrosaurus獨龍
`--Tyrannosaurinae 暴龍亞科=={暴龍>阿爾伯脫龍,后彎齒龍}=Deinodontinae恐齒龍亞科
| 未定:
| Deinodon恐齒龍
|--Alioramus分支龍
`--+-Albertosaurus阿爾伯脫龍
|--Gorgosaurus蛇發女怪龍
`--Tyrannosaurini 暴龍族=={暴龍>阿爾伯脫龍,蛇發女怪龍}
| 未定:
| ?Aublysodon后彎齒龍
| ?Shanshanosaurus鄯善龍
|--Daspletosaurus懼龍
|--Tarbosaurus特暴龍
`--Tyrannosaurus暴龍

暴龍超科

暴龍
侏羅紀時期暴龍類的化石資料殘缺不全,我們對其也知之甚少。發現於英國懷特島郎氏始暴龍(Eotyrannus lengi)相比髂鱷龍和斯托克斯龍等化石資料較多,約為整體骨骼的40%。科學家也較為確定始暴龍代表了代表了暴龍類一原始類型。始暴龍前上頜骨中的牙齒橫截面為類似字"D"的半圓形,其頭骨,肩部和後肢也與其他暴龍類相似。這具始暴龍個體長度約4.5米,其鼻腔骨骼癒合,但身體其他部位骨骼則沒有,顯示這具始暴龍可能是未成年個體。於後期其他暴龍不同的是,始暴龍手臂較長,具有三指。

1897年的暴風龍(現名傷龍)繪畫
正如前文中提到的,因為暴龍類是在肉食類恐龍全部滅絕後才演化出巨大的體型,填補了肉食龍類留下的空白,佔據食物鏈頂端。晚白堊世後半期北美洲暴龍類十分興盛,包括我們熟知的阿爾伯它龍,霸王龍等。阿爾伯它龍(Albertosaurus)發現於北美西北部的阿爾伯它,懷俄明,蒙大拿等地生活於約7千萬年前的晚白堊世,身長7-9米。阿爾伯它龍屬包括兩個種:肉食阿爾伯它龍(Albertosaurus sarcophagus)和大阿爾伯它龍(Albertosaurus grandis)阿爾伯它龍頭部粗壯,牙齒鋒利,向後彎曲,後肢長而強壯,善於奔跑,是十分強大兇猛的肉食恐龍。一些科學家推測阿爾伯它龍可能以小群體的方式捕獵,群體中包括少年和成年個體。
戈爾岡龍(Gorgosaurus)於阿爾伯它龍十分相近。戈爾岡龍同樣發現與北美西北部的阿爾伯它和蒙大拿,生活於7千5百萬年前,身長約8-9米,模式種為Gorgosaurus libratus。70年代科學家認為戈爾岡龍和阿爾伯它龍為同屬的動物,兩者之間的區別是由個體及生長差異造成的。根據命名原則,后建立的“戈爾岡龍屬”被取消,種被歸入阿爾伯它龍屬下,成為Albertosaurus libratus。但加拿大古生物學家Phillip Currie的近期研究結果表明戈爾岡龍具有的某些特徵與懼龍和霸王龍更接近,與阿爾伯它龍不是同屬生物。戈爾岡龍這一名稱得以保留下來。
懼龍(Daspletosaurus)於阿爾伯它龍的年代分佈一樣,但個體更大,長約9米,身體也較戈爾岡龍強壯很多,命名者根據這一特徵將懼龍的模式種命名為肌肉懼龍(Daspletosaurus torosus)意為肌肉發達的,可怕的蜥蜴。將懼龍和戈爾岡龍比較便可看出暴龍類一支的發展趨勢:身體增大變粗,頭骨同時增大,下頜更加巨大堅實,能夠附著更多的肌肉,咬力增大。
特暴龍(Tarbosaurus)發現於中國和蒙古,是晚白堊世末期東亞最巨大的肉食恐龍。確定的特暴龍種有兩個:艾氏特暴龍(Tarbosaurus efremovi)和勇士特暴龍(Tarbosaurus bataar)。已發現的特暴龍個體比霸王龍身體略小,勇士特暴龍身長9-12米;艾氏特暴龍更小,7-8米。特暴龍與北美洲的霸王龍十分相近,有些科學家甚至把他們劃分到一個屬,如Tyrannosaurus bataar。
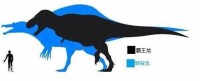
霸王龍和棘龍
最後值得一提的是發現於蒙古的分支龍(Alioramus),模式種遙遠分支龍(Alioramus remotus)。分支龍生活於6千5百萬年前的白堊紀晚期,與同時期其他暴龍類不同的是,分支龍保留了一些原始暴龍類的特徵。分支龍身體較小,5-6米長,口鼻部分十分細長而底平,長滿了椎形的骨質突起。一些科學家認為分支龍是其他大型暴龍類的幼年個體。但分支龍鼻腔中的氣室發育的很完善,是成年恐龍的特徵,因而派除了幼年個體的可能性。分支龍保留了暴龍類的原始特徵,包括細長而底平的口鼻部分等,明顯代表了暴龍類中一個演化分支。一些科學家同時指出,幼年的暴龍類顯示出祖先的原始特徵,因而頭骨形狀與分支龍類似。
6千5百萬年前一顆巨石從天而降,將本已開始衰敗的恐龍一掃而光。暴龍類也同鴨嘴龍,三角龍一起,永久的成為了歷史。
華麗羽王龍是由中國古生物學家在中國遼寧省西部早白堊世地層中發現一種新的暴龍類恐龍。它是中國遼西熱河生物群迄今發現的體型最大的恐龍之一。正如它的名字一樣,華麗羽王龍標本上保存了精美的羽毛印痕,這一發現為帶羽毛恐龍的家族再添一員。相關的研究成果發表在英國的《自然》雜誌上。根據目前發現最大的化石顯示,羽王龍體重1.4噸,體長8米,臀高2.5米。主要特徵包括大型的頭部、較瘦的軀體、中等長短的前肢、很長的腿、身體覆蓋著羽毛。
根據化石,研究人員預測:成年的華麗羽王龍體長在8米左右,頭骨長90厘米,體重大約1.4噸。雖然華麗羽王龍的體型比霸王龍小很多,但比遼西地區已知最大的帶羽毛恐龍意外北票龍還要大4倍。華麗羽王龍代表了已知體型最大的帶羽毛的恐龍。華麗羽王龍身上的羽毛只是非常簡單的絲狀物,代表了一種非常原始的羽毛類型;這種結構類似於小雞身上的絨毛,而與鳥類的體羽有所區別。華麗羽王龍龐大的體型和原始的絲狀羽毛表明其顯然不具有飛行能力,因此這些原始羽毛一個可能的重要功能是用來保溫。
值得一提的是:華麗羽王龍的體形與現在已知最大的帶羽毛恐龍:二連巨盜龍不相上下。可能在以後有望超過二連巨盜龍像鳥類的多樣性一樣,並不是所有羽毛恐龍都是輕重量動物,在1.25億年前現今中國境內生活著一種體型龐大的羽毛恐龍Yutyrannus huali,意思是“美麗的羽毛暴君”,它們的體長可達到8米長,臀高2.5米,體重達到1400千克。
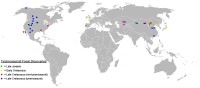
暴龍超科的化石只發現於北半球
科學家們通常將暴龍超科包含暴龍科與其最近親。但隨者種系發生學應用在古脊椎動物上,暴龍超科已有數個更明確的定義。在1998年,保羅·塞里諾(Paul Sereno)首次將暴龍超科作出親緣分支分類法的定義:跟暴龍具有更接近的共同祖先,而與新鳥亞綱具有更遙遠的共同祖先,這些物種所構成的基群演化支。在2004年,湯瑪斯荷茲(Thomas Holtz)提出一個較為狹窄的定義:親緣關係接近於暴龍,而離恐爪龍、似鳥龍、或是異特龍較遠的所有物種。2005年,塞里諾提出了一個新的定義:親緣關係接近於暴龍,而離似鳥龍、伶盜龍、以及傷齒龍較遠的所有物種。
髂鱷龍Iliosuchus -(中侏羅紀/英格蘭)
祖母暴龍 Aviatyrannis -(晚侏羅紀/葡萄牙)
冠龍 Guanlong -(晚侏羅紀/中國西部)
史托龍Stokesosaurus -(晚侏羅紀/西部美國)
帝龍 Dilong -(早白堊紀/中國東部)
小掠龍 Bagaraatan -(晚白堊紀/蒙古)
傷龍Dryptosaurus -(晚白堊紀/美國東部)
始暴龍 Eotyrannus -(早白堊紀/英格蘭)
獨龍 Alectrosaurus -(晚白堊紀/蒙古)
阿巴拉契亞龍 Appalachiosaurus -(晚白堊紀/美國東部)
嶼峽龍 Labocania -(晚白堊紀/墨西哥西部)
暴龍科Tyrannosauridae
艾伯塔龍 Albertosaurus -(晚白堊紀/美國西部)
分支龍 Alioramus -(晚白堊紀/蒙古)
懼龍 Daspletosaurus -(晚白堊紀/美國西部)
蛇發女怪龍 Gorgosaurus -(晚白堊紀/美國西部)
特暴龍 Tarbosaurus -(晚白堊紀/蒙古)
暴龍 Tyrannosaurus -(晚白堊紀/美國西部)
雖然古生物學家很早就以承認暴龍科的存在,但暴龍科的起源仍有很多爭論。在20世紀的大部分時間,暴龍科普遍被歸類於肉食龍下目,該下目包含了幾乎所有的大型獸腳類恐龍。肉食龍下目的異特龍科常被認為是暴龍科的祖先。在90年代早期,親緣分支分類法研究將暴龍科改列入虛骨龍類,這個理論早在20年代便有人提出,在90年代再度被提起。現在,暴龍科普遍被認為是種大型虛骨龍類。
在1994年,霍爾茲根據相同的踝部構造,例如第二、第四蹠骨在跗骨處接觸,把第三蹠骨夾住,將暴龍超科、單足龍科、似鳥龍下目、以及傷齒龍科,合稱為夾蹠龍類,是虛骨龍類的一個演化支。然而,基礎暴龍超科並沒有夾蹠型態的腳踝,例如帝龍,顯示這個特徵是平行演化而成的。夾蹠龍類已遭拆散,大部分古生物學家不再使用這名詞,而暴龍超科被認為是群基礎虛骨龍類,不屬於手盜龍形類。一個近年的研究提出虛骨龍科是暴龍超科的姐妹生物群,虛骨龍科包含虛骨龍與長臂獵龍(Tanycolagreus),生存於晚侏羅紀的北美洲。
以具有完整的化石而言,目前最基礎的暴龍超科是冠龍。其他的早期成員還有史托龍、祖母暴龍,兩者的化石較不完整。更著名的帝龍,被認為較冠龍、史托龍為衍化。傷龍過去難以分類,但數個近年的研究將它列為基礎暴龍超科,與暴龍科的關係較始暴龍、阿巴拉契亞龍為遠。獨龍出土於蒙古,較不著名,已確定是種暴龍超科,但它的演化位置仍不確定。其他被不同科學家認為可能是暴龍超科的物種,包括:小掠龍、嶼峽龍、以及被錯誤歸類于吉蘭泰龍的毛兒圖吉蘭泰龍(C. maortuensis)。發現於泰國早白堊紀地層的暹羅暴龍,最初被認為是種早期暴龍科,但現在普遍被認為屬於肉食龍下目。髂鱷龍的腸骨有個垂直棱脊,類似暴龍超科,可能是該超科的最早成員,但沒有足夠的化石可供確認。一個研究指出,某些美頜龍科的成員可能其實是基礎暴龍超科動物。
已知最早的暴龍超科動物出現在晚侏羅紀,包含:中國西北部的冠龍、美國西部的史托龍、以及葡萄牙的祖母暴龍。有些目前歸類於史托龍的化石,可能屬於祖母暴龍,這是因為該時期的北美洲與葡萄牙具有類似的動物群。髂鱷龍生存於中侏羅紀巴通階的英格蘭,如果髂鱷龍屬於暴龍超科,它將是該超科最早的成員,也意味暴龍超科的起源為歐洲。
早白堊紀的暴龍超科化石發現於北方各大陸。英格蘭的始暴龍與中國西北部的帝龍是這個時期唯二的暴龍超科成員,不過在猶他州雪松山組與日本的手取群發現了一些暴龍超科的前上顎骨牙齒,年代為早白堊紀。發現於中國內蒙古大水溝組的毛兒圖吉蘭泰龍(Chilantaisaurus maortuensis),有時被認為是早白堊紀的暴龍超科動物。
到了中白堊紀,歐洲已沒有暴龍超科的化石,顯示它們已在這地區滅絕。北美洲的達科他組,與哈薩克、塔吉克、烏茲別克的中白堊紀地層,都發現了暴龍超科的牙齒,以及可能的身體化石。年代最早且明確屬於暴龍科的化石,出土於北美洲與亞洲,年代為晚白堊紀的坎潘階。目前有兩個已承認亞科,艾伯塔龍亞科僅發現於北美洲,而暴龍亞科則分佈於北美洲與亞洲。阿拉斯加州曾發現過暴龍科的化石,該地過去可能是個陸橋,使兩大陸的動物可以散布。非暴龍科的暴龍超科動物,例如獨龍,可能還有小掠龍,與暴龍科共同生存於亞洲,但已在北美洲滅絕。在中白堊紀,北美洲東部被西部內陸海道所隔離開來。北美洲東部沒有發現過暴龍科化石,意味者暴龍科是在西部內陸海道產生后才演化出現;而基礎暴龍超科則在東部存活下來,例如傷龍與阿巴拉契亞龍,直到白堊紀末期。
中國遼寧省的早白堊紀地層義縣組,與鄰近的地層,已發現過許多虛骨龍類化石,保有長條纖維結構。這些纖維物通常被認為是原始羽毛,與有羽毛恐龍與鳥類的有分叉羽毛有相似處,但也有其他的假設存在者。在2004年,一個帝龍的化石發現了原始羽毛,這是第一個發現的暴龍超科羽毛痕迹。帝龍的原始羽毛具有分叉,類似現代鳥類的絨羽,但並非正羽,可能具有隔離體溫的功能。

懼龍的頭部與頸部
許多獸腳亞目恐龍的頭頂具有骨質冠飾,例如原角鼻龍與單脊龍,許多暴龍超科也擁有這個特徵。冠龍具有其中最複雜的頭冠,它們的鼻骨支撐一個大型冠飾,沿者頭顱骨中央直到后側。冠龍的頭冠上有數個大型洞孔,可減輕重量。帝龍的頭頂也有頭冠,較低矮、不明顯,由鼻骨與淚骨支撐者,沿者頭顱骨的兩側平行排列,在鼻孔後方往內彎曲,連接在一起,形成Y字形。暴龍科的閉合鼻骨通常表面不平。發現於蒙古的分支龍可能屬於暴龍科,鼻骨上有一排明顯的低矮骨質瘤,共五個;阿巴拉契亞龍的頭頂也有一排類似的骨質瘤,較為低矮,另外懼龍、艾伯塔龍、與特暴龍的某些標本也有類似的骨質瘤。艾伯塔龍、蛇發女怪龍、懼龍的眼睛前方,淚骨之上,具有明顯的眉角。特暴龍與暴龍也具有類似的隆起,但位置在眼睛後方,由眶后骨構成。
這些頭部裝飾物可能有展示物的功能,可能用來辨認物種或求偶。冠龍是一個“不利條件原理”(或稱“缺陷原則”)的例子,以掠食者而言,它們的大型、易碎頭冠,可能是獵食時的累贅。如果一隻具有易碎頭冠的動物身體健康,而且能夠成功獵食,這顯示該動物優於其他有較小頭冠的個體。冠龍的頭冠可能演化自性選擇,儘管獵食時造成麻煩,但偏重求偶上的功能,這點類似雄性孔雀的笨重尾巴,或是愛爾蘭麋鹿的大型鹿角。
在2001年的科學文獻里,曾提到發現暴龍類的剛孵化化石。
