RNA剪接
除去內含子將外顯子連接的過程
RNA剪接 (RNA splicing)是指從DNA模板鏈轉錄出的最初轉錄產物中除去內含子,並將外顯子連接起來形成一個連續的RNA分子的過程。RNA剪接機制的研究 ,是80年代生物化學和分子生物學領域中最有生機的研究課題之一,它不僅解決不連續基因轉錄產物的剪接問題,而且對於了解不連續基因的起源乃至整個生命起源與進化問題都是有力的推動,另外,核酸分子的催化功能的發現拓寬了人們對於酶的認識。
RNA剪接是真核細胞基因表達中非常重要的一個生物過程,通過RNA剪接,可以產生許多具有功能的,帶有編碼信息的mRNA,它對生物的發育及進化至關重要。所以RNA剪接識別是正確理解基因表達過程的重要一步,而剪接的識別的關鍵是依賴於剪接位點的判定,本文的工作即是針對剪接位點的正確識別進行研究。真核細胞pre-mRNA的剪接位點處存在一定的序列保守性,對於它所對應的cDNA序列而言,內含子5’端(供體位點)和3’端(受體位點)的鹼基幾乎都是GU和AG,因此稱為GU-AG規則。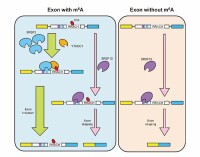
RNA剪接
RNA剪接可以有多種的方式。剪接的型式以內含子的結構及剪接所需的剪接因子而定。此外,RNA剪接還分為分子內 (intramolecular) 剪接 (cissplicing) 以及分子間 (intermolecular) 剪接 (transsplicing)。但不論哪一種途徑,移除的內含子都會被拋棄。
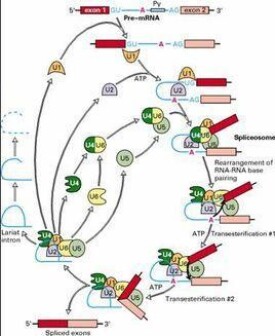
內含子經常存在於真核生物的蛋白質編碼基因(coding gene)中。在內含子里,需要有 5' 剪接位點(5' splice site)、3' 剪接位點(3' splice site)及剪接分枝位點(branch point)來進行剪接。剪接是由剪接體(Spliceosome)來催化,它是以五個不同的小核核糖核酸(snRNAs) 以及不下於一百個蛋白質所組成的大型核糖核酸蛋白質複合物,稱為小核核糖核蛋白(snRNP)。snRNP 的 RNA 會與內含子行雜交反應(hybridization),並且參與剪接的催化反應。
自剪接出現在稀少的內含子組成核酸酶,核酸酶在只有RNA的情況下代替了剪接體的功能。自剪接的內含子有兩種,稱為第I型及第Ⅱ型。第I型及第Ⅱ型內含子以與剪接體類似的方式進行剪接,但不需要任何蛋白質。這種相似性使人相信這些內含子與剪接體在演化過程上有著關連。自剪接亦可能是非常古老,且可能出現在一個還未有蛋白質的核糖核酸世界。雖然以下兩種剪接可以在沒有蛋白質的情況下進行,但依然會額外的使用5個RNA分子及超過50多個蛋白質,並水解多個三磷酸腺苷(ATP)分子。使用 ATP 是要提高剪接mRNA的準確性,避免出現錯誤。
以下兩次轉酯化是第I型內含子自剪接的特徵:
1.遊離鳥嘌呤核苷酸(被包在內含子中)的3'羥基,或是核苷酸輔助因子(即鳥苷單磷酸(GMP)、鳥苷二磷酸(GDP)、鳥苷三磷酸(GTP))攻擊內含子的5'剪接位點。內含子並不形成套索結構,而該鳥糞苷則會從內含子中轉移位置到內含子的5'位,從而成為第I型內含子的第一個核苷酸。
2.內含子5'剪接位點上游外顯子最後一個核苷酸的3'羥基變成親核基,而第二次交酯化/轉酯化會將兩個外顯子接合。
以下是第Ⅱ型內含子自剪接的特徵(與第I型相同是兩次交酯化):
1.內含子內特定腺苷的2'羥基攻擊5'剪接位點,從而形成一個套索。
2.5'外顯子的3'羥基新親核基於3'剪接位點引發第二次的交酯化/轉酯化反應,從而將兩個外顯子接合。
轉運RNA(tRNA)剪接是另一種較罕見的剪接方法,但是卻經常在 tRNA 出現。它的剪接反應涉及與剪接體或自剪接不同的生物化學過程。核糖核酸酶切開RNA,而連接酶(RNA ligase) 則將外顯子接合。這種剪接方式同樣不需要任何RNA分子來催化,而是一種全由蛋白質催化和作用的反應。整個過程中並未有交酯化/轉酯化作用。
RNA的剪接就是要把斷裂基因的轉錄本中的內含子除去。酵母細胞核中400個tRNA基因中約有40個是斷裂基因。這些基因均只有一個內含子,位於與反密碼子的3'側相隔一個核苷酸之處,長度為14至46bp。不同氨基酸的tRNA基因中的內含子不相同,因此,剪接酶類看來並不能識別任何共同順序。所有內含子中均有一段與tRNA的反密碼子互補的序列,因而使反密碼臂的構象發生了改變,即反密碼子被配對而使反密碼臂伸長了很多。在前體中僅反密碼臂受到影響,tRNA分子的其他部分仍保持其正常結構。
酵母tRNAphe中的內含子能與其反密碼子鹼基配對,從而改變了反密碼臂的結構。此剪切過程可分為兩個階段。第一步是磷酸二酯鍵的斷裂,這不需要ATP。這一步由一種內切核酸酶所催化。第二步是連接反應,需要ATP的存在,由RNA連接酶所催化。在無ATP時,產生的兩個tRNA半分子不能連接起來。這兩個半分子具有獨特的末端:其5'端有OH基,而3'有一個2',3'-環磷酸基。當加入ATP時,即發生第二步反應:兩個tRNA半分子先發生鹼基配對,形成成熟tRNA分子的構象,然後由RNA連接酶形成磷酸二酯鍵而將兩個半分子共價連接起來。2',3'-環磷酸基的存在並不限於酵母,在植物和哺乳動物的tRNA剪接反應中也有環狀基團的產生。在人的HeLa細胞中,RNA連接酶能將帶有2',3'-環磷酸基的RNA和另一帶有5'-OH基的RNA直接連接起來。酵母tRNA前體也可以在爪蟾的卵母細胞核提取液中正確地被剪接。這表示剪接反應沒有種屬特異性。爪蟾具有能識別酵母tRNA的內含子的酶類。
自身剪接反應
以前一直認為只有蛋白質有酶活性。這個概念在生物化學界已根深蒂固。然而近期發現RNA也可有酶活性。這種有酶活性的RNA有人稱之為ribozyme。
一種四膜蟲Tetrahumenathermophila的兩個主要rRNAs的基因和其他真核生物相類似(見前文),被轉錄在同一個初級轉錄本中。此轉錄本稱為35S前體RNA,較小的rRNA的序列在5'側,較大的rRNA(26S)序列則在3'側。在編碼26SrRNA序列存在一個單一的,短的(約400bp)內含子。如將這個35S前體RNA在體外溫育,可以發生自動剪接作用:內含子從前體中被切出,先呈線性RNA片段,後來又環化為環狀RNA。這個反應僅需要加入一種一價陽離子,一種二價陽離子,和一種鳥嘌呤核苷酸(G)。其他鹼基均不能代替G.但並不一定需要GTP;GDP;GMP和鳥苷都可以應用。這表示此反應並不需要能量供應。此外,此鳥嘌呤核苷酸必須有一個遊離的3'-OH基。這個G要連接到內含子的5'端上(通過通常的磷酸二酯鍵)。當線性的內含子成為環狀時,其3'端可連接在距離5'端15個核苷酸之處,從而將原來5'端和15個鹼基的節段(包括G在內)排除出去。這種反應基本上是一種磷酸酯轉移反應。外顯子A的3'-OH基可直接和外顯子B的5'端相連接。亦即一個磷酸酯可以直接被轉移到另一個上去,不需要經過中間步驟(如水解作用之類),因此磷酸酯鍵的能量被保存著。這解釋了為什麼此反應不需要水解ATP或GTP來供應能量。同時,兩次磷酸酯轉移反應似乎是緊密相連的,因為始終沒有找到過遊離的外顯子(A或B)。而線性內含子的環化則可以被看作是第三個磷酸酯轉移反應。在體外系統中進行剪接時,並不需要蛋白質的存在。RNA有能力自行剪接,故稱為自身催化作用(auto catalysis)。
T.Cech和S.Altman各自獨立地發現RNA具有催化作用。從而改變了生物催化劑的傳統概念。為此他們共同獲得了1989年Nobel化學獎。1978年Altman從純化的RNA酶P中分離出一種多肽和一種RNA(M1RNA)。最初的實驗結果表明,蛋白質和M1RNA單獨都沒有酶活性,但二者混合在一起又可恢復活性。其它生物材料的實驗結果表明M1RNA是RNA酶P活性所必須的。1983年Altoman證明,在較高濃度的Mg2+存在下,單獨的M1RNA就可以催化tRNA前體的成熟,而單獨的蛋白質則沒有這種能力。這樣,RNA即可看作是個酶。事實上,M1RNA的酶活性並不比RNA酶P的粗製品的活性低。原來認為蛋白質賦予酶的活性,RNA只起某種輔助作用(例如幫助蛋白質與其底物結合),但現在發現這兩種功能已經倒轉。Cech給具有催化活性的RNA定名為ribozyme。
很長時間以來,人們就試圖自己設計和生產酶分子,但因蛋白質分子結構上的複雜性,迄今為止,尚無成功的例子。近年來隨著ribozyme的發現,人工酶(新概念下的酶,它的構件分子是核苷酸)的設計又產生了新的希望。澳大利亞的科學家就設計了九個ribozyme分子,它們都具備內切酶的活性,且切割位點有高度特異性。同時,ribozyme的活性隨pH、溫度、及陽離子濃度的變化而變化,顯示出典型的酶特性。由於ribozyme的作用位點高度特異,故可以用來切割特定的基因轉錄產物(RNA)。有人將這種切割作用叫做抗基因活性。因為切割的結果破壞了RNA,也就是抑制了基因的表達。這種特性為我們進行基因和病毒的治療提供了一個可行的途徑。
某些線粒體中的內含子也是自身剪接的內含子。一些常見的真菌,如粗糙脈孢菌(Neurosporacrassa),酒酵母(Saccharomycescerevisiae)等的線粒體內含子都能進行自身剪接,像四膜蟲中進行的磷酸酯轉移反應一樣。
能編碼蛋白質的內含子
某些真菌線粒體中的內含子有很不尋常的結構,這些內含子有編碼順序,這些順序的翻譯對於該順序所在的內含子的剪接是必須的。編碼細胞色素b的box基因在僅剪去內含子1時,會產生RNA成熟酶的mRNA。當內含子2亦被剪去時,才是細胞色素b的編碼順序的開端。線粒體中的box基因是編碼細胞色素b(cytb)的。box基因中的外顯子1有417bp,編碼cytb的N端139個密碼子。內含子1有765bp,不編碼。外顯子2非常短,僅有5個密碼子。其後是一個很長的內含子2。內含子2的特點是其開頭840bp是一個開放讀框(ORF),有280個密碼子,其最後一個密碼子為終止密碼子。當將內含子1剪去后,翻譯即從外顯子1起始,通讀過外顯子2而進入內含子2的ORF,產生一個有423個氨基酸殘基的蛋白質(其中144個是cytb的N端氨基酸,279個則由內含子2編碼,稱為RNA成熟酶(maturase)。RNA成熟酶是特異地用來剪去內含子2的。這樣,這個酶促反應便成為一個非常敏感的負反饋徑路。去除內含子2后,外顯子1和2即與外顯子3相連接,因而破壞了編碼成熟酶的順序。
細胞核中的剪接連接點
剪接連接點(splicing junctions)是指在切斷和重接位點處的兩旁的順序。在內含子左側的連接點稱為供體(donor),在內含子右側的稱為受體(acceptor)。在細胞核的結構基因(即編碼多肽的基因)中的所有內含子在外顯子-內含子連接處均有GU...AG的共同順序。較詳細的共同順序如下,供體位點受體位點:
外顯子...AG↓GUAAGT...內含子...Py10CAG↓...外顯子
箭頭表示切斷的鍵。這些還是較短的共同順序,存在於幾乎所有的真核生物中。
從上述共同順序可見供體和受體位點之間並無互補現象,所以不可能想像這兩個位點會通過鹼基配對而結合一起,以便於內含子的切除。正確的剪接並依賴於天然前體RNA分子的完整性。一個外源基因如果存在於病毒順序中,仍能很好地被剪接。另外,前體RNA亦能在不同的組織,或甚至不同物種的細胞中被正確地剪接。這都表示剪接作用是很保守的。一個真正基因中一個外顯子可以和另一個基因的外顯子連接起來。例如,將SV40(猴病毒40)的早期轉錄單位的第一外顯子和小鼠β珠蛋白的第三外顯子相連,這樣形成的雜交內含子仍能正確地被剪接。即SV40的內含子的供體位點(1I)可以剪接到小鼠β珠蛋白的內含子的受體位點(r2)上。
套索的產生
HeLa細胞的核提取液能夠剪接純化的RNA前體,這表示剪接作用並不與轉錄作用相關連。RNA的修飾也和剪接無關,例如珠蛋白RNAs即使缺少poly(A)尾鏈,也沒有加帽,仍能正常地被剪接。剪接過程可分為兩個階段,與上文所述自身剪接不需能量不同,核內剪接需要用ATP。
在第一階段中,內含子左端(供體位點)處被切斷,形成兩個分離的RNA分子,即左外顯子和右內含子-外顯子。左外顯子此時為-線性分子,但右內含子-外顯子則不然:內含子左端(5'端)以5'-2'鍵與在內含子右端上游約30鹼基處的CUGAC序列(共同序列)中的A相連接,於是形成一個"套索"(lariat)"。在第二階段,在受體位點處被切斷而將此套索狀的內含子剪去;同時分離的右外顯子即與左外顯子相連接。套索然後被"脫支(debranch)"而形成一線性內含子。在這種剪接機構中,有三個很短的共同順序,即供體和受體位點處於一個和套索分支處的一個序列。用酵母做的實驗證明:分支處的共同順序如果發生突變或缺失將使剪接不能進行。這個共同順序常稱為UACUAAC盒。它和左側的供體位點共同順序互補,但研究證明兩者並不發生鹼基配對。在高等真核生物中此分支靶順序的保守性較小。
snRNAs的作用
真核細胞有細胞核和細胞漿中都含有許多小RNA,它們約有100到300個鹼基,每個細胞中可含有105-106個這種RNA分子。它們是由RNA聚合酶Ⅱ或Ⅲ所合成的,其中某些像mRNA一樣可被加帽。在細胞核中的小RNA稱為snRNA,而在細胞漿中的稱為scRNA。但在天然狀態下它們均與蛋白質相結合,故分別稱為snRNP和scRNP。某些snRNPs和剪接作用有密切關係。有些snRNPs分別和供體及受體剪接位點以及分支順序相互補。snRNAs中最受注意的一個是U1,它普遍存在於哺乳動物、鳥類和昆蟲細胞中。人U1snRNP中除RNA外還8個蛋白質分子。人U1snRNA的可能二級結構如圖。其5'端的11個核苷酸是單鏈的,並有一段和內含子左側的供體序列互補。在供體位點處的互補序列通常為4-6bp。在體外,完整的U1snRNP粒子能和左側共同順序結合,但純化的U1snRNA卻不能。U1snRNA參與剪接的證據是抗U1snRNP的抗體可以在體外抑制剪接作用;而且如果從系統中將U1snRNP除去,剪接即不能進行。事實上,除去U1sn RNA5'端的幾個核苷酸即可抑制體外剪接作用。
有可能U5snRNA能識別右側(受體位點)共同順序;而U2snRNA,則具有與分支位點互補的順序。抗U2snRNP的抗體可以和U2snRNA及包括分支位點在內的內含子所形成的複合體發生免疫沉澱。U1和U2可能參與剪接反應的起始階段,因為U1和U2的滅活將阻止左側連接點的切斷和套索的形成。另外兩個snRNAsns(U4和U6)可能亦參與剪接作用,但它們的功能尚不詳。一般認為,脊椎動物細胞有6種不同的snRNAs,稱為U1、U2、U3、U4、U5和U6。最小的是U6,有約100核苷酸長。最大的是U3,也不過215核苷酸長。它們和蛋白質結合成snRNPs。系統性紅斑狼瘡(SLE)患者和某些風濕病患者的血清中常可檢出對snRNPs中某些蛋白質的自身抗抗體。
核內不均一RNA
編碼蛋白質的結構基因是在核漿中被轉錄的。但是核漿中的RNA卻並不像mRNA。核漿RNA要大得多,很不穩定,並且其順序的複雜性也要大得多。由於它的大小很不一致,故稱核內不均一RNA(hnRNA)。已經證明mRNA確實是從hnRNA生成的。細胞漿內的mRNA平均只有1800-2000個鹼基。而哺乳動物的hnRNA平均有8000-10000個鹼基,其範圍很廣泛,從2000-14000鹼基均有,所以一般要比mRNA大4-5倍。如果以5倍計算,由於正常哺乳動物細胞中測得mRNA僅佔hnRNA量的5%,則相當於有25%的hnRNA可轉變為mRNA。這意味著有3/4的hnRNA即在核內降解。hnRNA被切除內含子后即成為mRNA,並進入細胞漿內。但在切除內含子之前,hnRNA可先加帽和加poly(A)尾鏈。有一種稱為poly(A)聚合酶的酶可以用ATP為底物,以加上poly(A)尾鏈,這是hnRNA轉變為成熟的mRNA所必須的。在哺乳動物細胞中,僅約30%的hnRNA被多聚腺苷酸化,而mRNA中卻約70%是有poly(A)尾鏈的。可能多聚腺苷酸化是hnRNA分子要被加工的信號。hnRNA的尾端要被切去一段,然後才加上poly(A)。因為RNA聚合酶在轉錄時即已通過了相當於加上poly(A)的位點,故hnRNA尾端的多餘部分要由內切核酸酶切去,才能加上poly(A)。
RNA的編輯是某些RNA,特別是mRNA前體的一種加工方式,如插入、刪除或取代一些核苷酸殘基。介導RNA編輯的機制有兩種:
特異性脫氨基作用
比如:哺乳動物的載脂蛋白mRNA的編輯 其蛋白編碼區的DNA序列在所有組織中都一樣;在肝臟中該基因轉錄為完整的蛋白質,而在腸中合成的Pr長度只有其一半(只是全長載脂蛋白的N端),是由於2153位上的密碼子從CAA突變為UAA(使編碼谷氨醯胺的密碼子變為終止密碼子)。還有一個例子 大鼠腦中的谷氨酸受體蛋白mRNA經編輯后,分子中有多個編碼谷氨酸的密碼子變成了在控制通過神經遞質的離子流過程中又主要作用的精氨酸,表明RNA的編輯可能是充分發揮生理功能必需的。
以上兩種情況分別由胞嘧啶和腺嘌呤脫氨酶所催化;通常情況下該酶促反應的特異性不強,腺嘌呤脫氨酶可作用於雙鏈RNA區的任何腺苷酸殘基;但是,RNA的編輯發生在帶有具催化作用的脫氨酶亞基的複合體中,有附加的RNA結合區能幫助識別所編輯的特異性靶位點。
尿嘧啶插入或刪除
指導RNA:特異性插入這些殘基的信息來自因為它含有與編輯后mRNA互補的核苷酸序列,指導RNA與被編輯區及周圍部分核酸序列雖然有相當程度的互補性,但該RNA上存在一些未能配對的腺嘌lin,形成缺口為插入尿嘧啶提供了模板。
RNA編輯具有重要的生物學意義:
校正作用;調控翻譯;擴充遺傳信息
mRNA在某些情況下不是以固定的方式翻譯,而可以改變原來的編碼信息,以不同的方式進行編碼 包括:+1/-1移位;核糖體跳躍
六大類化學修飾:甲基化;去氨基化,硫代(S代替鹼基的O分子),鹼基的同分異構化(尿嘧啶變構生成假尿嘧啶);二價鍵的飽和化;核苷酸的替代(用不常見的核苷酸替換常見的核苷酸)核酶
核酶(ribozyme)是指一類具有催化功能的RNA分子,通過催化靶位點RNA鏈中磷酸二酯鍵的斷裂,特異性地剪切底物RNA分子,從而阻斷基因的表達.
核酶的催化功能與其空間結構有密切關係,目前已知有多種特殊結構的核酶:
RNaseP的RNA鹼基(M1 RNA)、鎚頭型、髮夾型丁型肝炎病毒RNA、1類內含子和2類內含子,大多有hammerhead structure。
不同的核酶可分為兩類:
1 剪切型核酶:只剪不接,如M1 RNA
2 剪接型核酶:該酶具有序列特異的內切核酸酶、RNA連接酶等多種酶活性,如1類和2類內含子
為了識別在RNA干涉(RNAi)和微RNA介導的基因表達調控中所涉及的因素,Gary Ruvkun及其同事對86種真核生物進行了系統發生分析,所得到的候選物再用轉錄和蛋白組相互作用數據進行Bayesian分析,來估計它們參與小RNA調控的概率。所識別出的小RNA輔因子中大約一半是RNAi沉默所必需的,其他當中的很多參與剪接,說明在這兩個過程之間存在一個聯繫。
RNA剪接及其機制的研究,不僅解決了不連續基因“連續”轉錄產物的問題,而且對於了解不連續基因的起源乃至整個生命起源與進化等問題,均產生極大的推動作用,另外,由此發現了核酸分子的催化功能,進一步拓寬了對於酶的認識。不連續基因中的介入序列稱為內含子;被內含子隔開的基因序列稱為外顯子(exon)。一個基因的外顯子和內含子都轉錄在一條原初轉錄物RNA分子中,然後把內含子切除而把外顯子連接起來,才能產生成熟的RNA分子,這個過程叫做RNA剪接(RNA splicing)。內含子上遊方向的一個剪接點稱為5’剪接點或左剪接點,內含子下遊方向的一個剪接稱為3’剪接點或右剪接點。Davies等人於1982年設定了“中部核心結構”(central structure)在內含子剪接中的作用,組成這種中部核心結構的是4個10~12鹼基的保守序列,並構成相應的二級結構。但並非所有的內含子都含有這樣的二級結構和保守序列 。
(一)第1類內含子自我剪接-rRNA的自我剪接
第1類內含子,其5’剪接點和3’剪接點的序列絕大部分為…外顯子…U↓…內含子…G↓…外顯子…,除了剪接點序列特徵之外,第1類內含子還具有比較保守的4種10一12核苷酸的序列,分別以5’-P-Q-R-S-3’表示,P、Q、R、S的一致序列。序列能與Q序列互補,R序列能與S序列互補,形成一個所謂中部核心結構。位於內含子序列中靠近5’剪接點的一段序列與兩個剪接點的邊界序列配對,將兩個剪接點拉在一起便於磷酸二酯鍵的切斷和再連接。內含子中能與兩個剪接點邊界序列配對的一段序列稱為內部引導序列(intemal guide sequence,IGS)。如內部引導序列GGACGG,與其5’剪接點上游的外顯子3’末端區的序列配對而形成雙鏈結構。此外,內含子形成了包括內部引導序列在內的9個雙鏈結構區 。
(二)第Ⅱ類內含子的自我剪接
第Ⅱ類內含子,其5’剪接點和3’剪接點的序列多為…外顯子…↓GUGCG…內含子…嘧啶鹼AU↓…外顯子…,除了剪接點序列特徵之外,在離3’剪接點上游6-12bp有一段比較保守的序列,一致序列為CUCAC,在這一保守序列A的兩側各有一段3~5核苷酸的短序列能與上遊方向的核苷酸互補,而A總是不包含在這些互補的序列之中,也就是說,在形成部分雙鏈結構中,A總是處於向外突出的“芽狀物”之中,而且該A啟動第一次轉酯反應。此外,內含子含有6個雙鏈結構區,內含子5’端形成的雙鏈結構區I含有兩個外顯子結合位點,可以與5’剪接點上游的外顯子3’末端區的序列配對而形成雙鏈結構 。
(三)第Ⅲ類內含子的剪接-hnRNA的剪接
核基因hnRNA內含子的剪接點序列為…外顯子…↓GU…內含子…AG↓…外顯子…,這就是普遍適用的所謂Breathnach-Chambon規則(GU-AG規則)(GU-AG rule),此規律不適合於線粒體和葉綠體的內含子,也不適合於tRNA和某些編碼rRNA的核結構基因,酵母的分支位點序列是高度保守的,其共有序列為UAC-UAAC。剪接點序列和分支位點保守序列在hnRNA剪接中非常重要,如果這兩個保守序列突變就不能正確的剪接hnRNA。然而,單靠這樣簡單的保守序列是不可能做到hnRNA正確剪接,snRNA參與了這種作用 。
(四)第Ⅳ類內含子的剪接-tRNA的剪接
酵母基因組共有約400個tRNA基因,含有內含子的基因僅佔十分之一。內含子的長度從14到46個鹼基對不等,它們之間並無保守序列,切除內含子的酶識別僅是共同的二級結構,而不是共同的序列。通常內含子插入到靠近反密碼子處,與反密碼子鹼基配對,未成熟tRNA的反密碼子環不存在,而是以插入的內含子所構成的環 。
(五)順式和反式剪接
內含子剪接一般都是發生在同一個基因內,切除內含子,相鄰的外顯子彼此連接,這種剪接稱為順式剪接( cis-splicing),但也有另一種情況,即不同基因的外顯子剪接,這種剪接稱為反式剪接(trans-splicing) 。
可變剪接(alternative splicing)是指在同一個mRNA前體內部數個外顯子之間產生的差異性連接。這種剪接可以使同一個基因在不同的發育階段、不同分化狀態甚至不同生理狀態下,得到多個相似但有差異的mRNA,進而被翻譯為氨基酸序列相近似、性質和功能有差異的蛋白質。高度通用性的剪接位點GU-AG被同樣高度通用性的剪接體成對識別是剪接過程發生的先決條件 。
可變剪接暗示了內含子的一種功能,即:由於內含子的存在,使可變剪接成為可能,其意義在於可使一個基因表達出多種蛋白質,擴大了DNA中遺傳信息的含量 。
基本信息
- 中文名
- RNA剪接
- 外文名
- RNA splicing
- 目標
- 除去內含子
- 方式
- 多種
- 意義
- 對生物的發育及進化至關重要
- 學科
- 生命科學