翻譯
生物學術語
翻譯是蛋白質生物合成(基因表達中的一部分,基因表達還包括轉錄)過程中的第二步(轉錄為第一步),翻譯是根據遺傳密碼的中心法則,將成熟的信使RNA分子(由DNA通過轉錄而生成)中“鹼基的排列順序”(核苷酸序列)解碼,並生成對應的特定氨基酸序列的過程。但也有許多轉錄生成的RNA,如轉運RNA(tRNA)、核糖體RNA(rRNA)和小核RNA(snRNA)等並不被翻譯為氨基酸序列。
翻譯是中心法則中一個不可或缺的過程,對生物機體的性能有著不可或缺的作用。
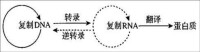
中心法則
mRNA是翻譯的模板。在原核生物和真核生物細胞內,mRNA的化學基礎有所差異。
原核生物mRNA
在原核細胞內,參與翻譯的mRNA具有以下特點:
(1)具有多個開放閱讀框(ORF),即多順反子,意味著同一條mRNA可以編碼多個蛋白。特別注意可讀框之間不重疊(除移碼翻譯涉及終止密碼子和起始密碼子的2個鹼基重疊)。
(2)具有較為保守的核糖體結合位點(RBS)GGAGG,位置大概在起始密碼子上游的3~9個鹼基。
(3)自身可通過RBS招募小亞基核糖體RNA(16S·rRNA)。
(4)存在移碼編譯。在一些情況下,第一個ORF的終止密碼子UGA和第二個ORF的AUG重疊,形成一個序列AUGA。當核糖體遇到UGA終止翻譯后可以隨即重啟,向後移動-1位至起始密碼子AUG,開始第二個ORF的翻譯。如此的生物學意義是當且僅當第一個ORF成功翻譯后,才會開啟第二個ORF的翻譯。
真核生物mRNA
相比原核細胞mRNA,真核細胞內參與翻譯的mRNA具有以下不同:
(1)總是單ORF的(即每條鏈只能編碼一個蛋白),即單順反子。
(2)沒有核糖體結合位點(僅有部分含有較為保守的Kozak序列:G/A——AUGG,其功能尚不完全明確)。
(3)核糖體的招募需要5'端的特殊結構(5'端加帽),加帽的反應過程可以簡要表示如下:
PPP-→PP-+Pi
PP-+GMP→G-PPP-
總式:PPP-+GMP→G-PPP-+Pi
(4)起始因子結合5'端帽子結構(詳見翻譯的起始)以招募核糖體。
(5)核糖體從5'端開始掃描,直到遇到AUG開始翻譯(以往認為大多是遇到第一個AUG即開始翻譯,後續觀點認為這種情況並不常見,具體機制尚不完全明確)。
tRNA的二級結構如圖1所示,其在原核生物和真核生物均相對保守。主要結構有D-loop(D環)、T(C)-loop(T環)、Anticodon-loop(反密碼子環)、Accepter Arm(受體臂)、3'端CCA保守序列、Discriminator(識別鹼基)、Variable-loop(可變環)等。
由於翻譯的氨基酸需要,tRNA在單個細胞內可有很多種,下面將討論各種mRNA之間的保守部分和可變部分。
保守部分
在不同的tRNA中,以下部分是所有tRNA共有不變的:
(1)3'端CCA序列(由相應酶不依賴特異性模板而添加的)。
(2)保守鹼基,包括:位於T-loop的7個、位於D-loop的4個、位於Anticodon-loop的1個)
(3)位於各臂間的多個鹼基對。
可變部分
在不同的tRNA中,以下部分是特異性的:
(1)反密碼子。
(2)Variable-loop(可變環)的大小:1類在3~5個鹼基對左右,2類在13~21個鹼基對左右。
(3)識別子(位於3'端的四個鹼基),其主要用於區分不同種類的tRNA。
鹼基配對原則
翻譯過程嚴格按照鹼基互補配對原則進行。值得注意的是,轉錄組成RNA的鹼基中,沒有T(胸腺嘧啶),而是U(尿嘧啶),所以配對原則如下所示。
DNA -> RNA
A ,腺嘌呤-> U,尿嘧啶
T ,胸腺嘧啶-> A ,腺嘌呤
G ,鳥嘌呤-> C,胞嘧啶
C,胞嘧啶-> G,鳥嘌呤.
由於密碼子的擺動性,反密碼子第1位鹼基與密碼子第3三位鹼基之間配對稱多樣性,可能配對方式如下:
I ->A,U,C
U ->A,G
G ->C,U
其中,I代表次黃嘌呤核苷(inosine),在tRNA反密碼子中出現。而其他兩位的配對方式即是嚴格的沃森-克里克配對方式。
特彆強調的是,雖然氨基酸種類很多,但是由於tRNA密碼子的擺動性,可以縮減所需要的tRNA數量。比如異亮氨酸的密碼子是AUA、AUC或AUU,則對應的反密碼子只需要是UAI一種即可。
翻譯的化學本質是單個氨基酸脫水縮合形成肽鏈,這一過程需要多種酶的參與。而在體內,多種酶參與的多種化學反應組成了翻譯的生物化學途徑。就化學層面來看,翻譯主要涉及到三個化學步驟:氨基酸的腺苷化(Amino Acid Adenylation)、tRNA裝載(tRNA charging)、肽鍵的形成。
腺苷化反應和tRNA裝載
這一過程是ATP和氨基酸反應最終生成2Pi、氨基酸(AA)-AMP的過程。氨基酸的羥基中的氧原子除了和C、H原子分別形成共價鍵之外,還具有獨立的2個電子對。由氨基酸的羥基向三磷酸腺苷的第一個磷酸發起攻擊,在保留第一個高能磷酸鍵的前提下,使PPi脫離,隨後形成2Pi。
這個化學過程可簡要概括如下:
AA + ATP → PPi + AA-AMP
PPi → 2Pi
總式:AA + ATP → 2PPi + AA-AMP
特別值得注意的是,在這個反應中,三磷酸腺苷內的高能磷酸鍵得以保留。
tRNA裝載過程是AA-AMP和tRNA發生反應最終形成AA-tRNA的過程。其化學本質是tRNA受體臂上的3'端羥基-OH攻擊AA-AMP上連接P原子和C原子的O原子,最終形成裝載完畢的tRNA。通過這種方式裝載完畢的tRNA依然保留了一個高能鍵。高能鍵的保留對於後續反應的發生相當重要。
這兩部反應是由氨醯-tRNA合成酶(AA-tRNA synthetase)催化完成的。後續將對該酶的結構生物學基礎做進一步探討。
肽鍵的形成
這一步反應是整個分子生物學過程的核心,但其化學本質很簡單,重點是其生物體內催化的過程。在以往的觀點裡,核糖體rRNA的具體序列或許對於肽鍵形成至關重要,因為在核糖體的反應核心並沒有蛋白質的參與,提示著rRNA對於肽鍵的合成起到主要的催化作用。而經過後續研究,當前普遍認為rRNA對於核心反應的催化並非具體序列依賴的化學催化,而更像是整體序列共同作用導致的物理結構催化。即rRNA通過整體序列(和蛋白互作)形成特定的結構,確保核糖體A位點和P位點的;兩個tRNA受體臂上的氨基酸能夠近距離正確接觸,催化反應的發生。
而tRNA自身的結構,甚至是氨基酸本身其他結構都在這個反應里起到了一定程度的重要催化作用,來確保這個反應的發生。
參與翻譯生化反應的有多種酶,但其核心生化反應主要由兩類酶參與:催化腺苷化反應和tRNA裝載的氨醯-tRNA合成酶、催化肽鍵合成的核糖體核酶。下面將進一步探討這兩種酶的結構生物學基礎,以及它們確保反應準確發生的校正機制。
氨醯-tRNA合成酶
氨醯-tRNA合成酶有四個結構域和三個活性位點。由於每種tRNA只能結合特定氨基酸,所以氨醯-tRNA合成酶必須確保tRNA和氨基酸之間的正確配對。
其四個結構域分別結合tRNA受體臂(第1結構域)、反密碼子區域(第2結構域,其中1個鹼基用來被識別)、ATP和正確AA(第3結構域)、錯誤AA(第4結構域)。
三個活性位點分別位於第1結構域(催化tRNA裝載)、第3結構域(催化腺苷化反應)、第4結構域(催化錯誤氨基酸水解)。
錯誤氨基酸在這裡一般指的是和該tRNA不匹配,但又由於化學性質類似而容易被錯誤裝配的氨基酸。氨醯-tRNA第3結構域內的AA結合位點對於匹配的正確AA具有高親和力,而對錯誤AA親和力較低,但依然會有概率結合而催化後續的兩步反應。但由於第4結構域對錯誤AA的高親和力,可以大概率將周圍的錯誤AA先行“捕捉”並且水解,防止其參與後續反應。這就是氨醯-tRNA合成酶的第一個校正機制,是化學或酶學機制。
第二個校正機制是動力學機制。正確的AA和tRNA組由於在合成酶中處於正確的位置而更容易得以被催化,導致反應的發生。
值得注意的是,翻譯和轉錄的校正機制往往沒有基因複製的高保真度。從生物學意義角度來看,翻譯的單個氨基酸出錯只會影響一個蛋白的功能,而基因複製錯誤則可能會影響一類蛋白終生的表達。
核糖體核酶
核糖體由大小兩個亞基組成,其rRNA佔到組分的50%,剩餘的50%是一些小型蛋白。蛋白質的主要作用是維持rRNA的正確摺疊,但值得注意的是,所有的催化作用都是rRNA介導的。
核糖體有兩個通道:mRNA-tRNA通道(貫穿三個tRNA結合位點:A、P、E)和肽鏈出口通道。大部分新合成的肽鏈都是在離開核糖體后才開始摺疊,而在肽鏈出口通道可以先行形成α螺旋。
核糖體的三個位點各自結合不同的組分。A位點結合AA-tRNA,P位點結合肽鏈-tRNA和起始AA-tRNA。E位點結合即將釋放的tRNA。最重要的肽鍵合成發生於A位點和P位點之間,故位於這兩個位點的tRNA的受體臂3'端在空間結構上距離很近以方便反應的發生,而E位點則距離較遠。在A、P位點的tRNA均和mRNA存在氫鍵互作,而在E位點的tRNA距離mRNA較遠,並無互作,方便其儘快脫離。
在核糖體中,有兩個重要的區域:肽醯轉移中心(肽鍵合成)和解碼中心(mRNA-tRNA互作)。在tRNA和rRNA的共同作用下,肽醯轉移得以催化。而tRNA的正確匹配在肽鍵合成過程中是必須的(詳見下文翻譯的延伸)。
(一)原核細胞
原核細胞的翻譯起始過程大概可以分為以下幾個過程:
(1)翻譯起始因子IF3結合到小亞基的E位點,同時也橫跨至P位點;(這一過程在起始之初就已經完成)起始因子IF1結合至A位點;
(2)起始因子IF2·GTP被IF3和IF1招募至P位點;
(3)起始fMet·tRNA一方面被mRNA起始密碼子AUG招募,另一方面被已經結合到P位點的IF2·GTP招募,進一步結合到P位點,這一過程伴隨著核糖體小亞基通過mRNA上的核糖體結合位點序列(RBS)識別結合到mRNA上;
(4)當tRNA和mRNA配對后,起始因子IF3穩定性降低;
(5)IF2·GTP進一步招募大亞基;
(6)大亞基的結合催化IF2上GTP的水解,形成對P位點親和力較低的IF2·GDP,後者隨後從核糖體上脫離;
(7)由於IF2的脫離,大小亞基結合進一步緊密,置換出IF3和IF1,形成了起始狀態的核糖體複合物。
註:實驗分析表明,當IF3和IF1不存在時,起始tRNA傾向於在IF2之前結合至P位點,提示起始tRNA可以直接被起始密碼子招募;當IF3和IF1存在時,起始tRNA傾向於在IF2之後結合P位點,提示IF2可以被IF3和IF1有力招募,而tRNA也可以進一步被IF2招募。
(二)真核細胞
真核細胞會涉及到更多的起始因子包括:eIF1,1A,3,5,4(EGAB),2,5B,1A。其中它們和原核細胞起始因子按照功能可以大體劃分配對如下:
| 真核細胞 | 原核細胞 |
| eIF1,eIF3,eIF5 | IF3 |
| eIF2 | IF2 |
| eIF5B | |
| eIF4E,eIF4G,eIF4A,eIF4B | — |
| eIF1A | IF1 |
以核糖體為核心,真核細胞翻譯起始過程也可概括成如下步驟:
A. 核糖體的前期準備
(1)eIF1,3,5圍繞E位點結合至小亞基,eIF1A圍繞A位點結合至小亞基;
(2)eIF2·GTP在胞質中結合Met-tRNA形成三原複合物;
(3)三原複合物進一步結合到小亞基複合物(小亞基以及eIF1,1A,3,5)中小亞基P位點上形成43S複合物;
B. mRNA的準備
(1)eIF4E識別mRNA的5‘端帽子;
(2)eIF4G作為支架蛋白結合至eIF4E並連接eIF4A,eIF4A結合至mRNA上;
(3)eIF4B和eIF4A互作,激活eIF4A的活性;
(4)eIF4A利用其解旋酶活性解旋mRNA,去除其二級、三級結構;
C. mRNA的裝載
(1)43S複合物通過eIF3和eIF4E互作,eIF1A和eIF4A互作將mRNA進行招募形成48S複合物;
(2)eIF4A利用其解旋酶活性,充當轉位酶作用,移動mRNA進行掃描;
(3)遇到第一個起始密碼子AUG后,密碼子和tRNA配對,阻礙了eIF4A的轉位,掃描停止;
(4)二者的配對,刺激eIF2的GTP水解,eIF2·GDP和小亞基親和力較低,從小亞基複合物上脫離下來,伴隨著其他除eIF4A以外的起始因子全部脫離;
(5)eIF5B·GTP被起始tRNA和eIF1A招募;
D. 大亞基的裝載
(1)eIF5·GTP招募大亞基;
(2)大亞基的結合刺激eIF5上的GTP水解,eIF5·GDP親和力較低,從核糖體上脫離下來;
(3)大亞基獲得更高親和力,進一步將eIF1A置換出來,進入起始狀態。
註:並非所有真核細胞內的翻譯均需要eIF4E對5’端帽子的識別,有下列三種特殊情況利用內部核糖體插入位點(IREs)進行RNA的裝配:
| Group 1 | 利用特殊結構直接招募核糖體大小亞基,不需要任何起始因子和起始tRNA。 例:一些病毒如CrPV入侵真核細胞后,通過抑制起始因子進一步抑制真核細胞內部的蛋白表達,但其自身特殊的mRNA結構可以直接招募大小亞基來繞過起始因子和起始tRNA階段,直接進入翻譯的中間延伸狀態,獲得核糖體翻譯的絕對選擇性。 |
| Group 2 | IREs招募小亞基,但依然需要部分eIF和起始tRNA。 例:部分IREs直接結合eIF4G進一步招募eIF4A。 |
| Group 3 | 需要IREs特殊的結合蛋白,同時也需要大部分eIF和起始tRNA。 例:細胞凋亡。細胞在凋亡時,希望減少其他蛋白的表達(通過抑制eIF4E),而大量表達凋亡相關蛋白。如死亡相關蛋白5(DAP5)利用其IREs序列招募相關因子(ITAF),後者充當eIF4G的eIF4A結合區域的作用,繞過eIF4E,直接結合eIF4A進行後續裝載。通過這樣的機制可以巧妙的降低其他蛋白(需要eIF4E)的表達,但是提高凋亡相關蛋白的表達。 |
此過程在真核細胞和原核細胞中高度類似,下面只以原核細胞為例進行討論。涉及到的因子主要有EF·Tu和EF·G,在真核細胞中對應的名稱分別是是eEF1和eEF2。
A. tRNA的轉運和入位
(1)非起始AA·tRNA結合EF·Tu·GTP形成一個三元複合物;
(2)該三元複合物結合至核糖體P位點,tRNA反密碼子和mRNA密碼子進行配對,伴隨著tRNA構象改變;
(3)小亞基RNA中G530、A1492、A1493等位點和密碼子及反密碼子互作,使其穩定配對;
(4)EF·Tu在核糖體大亞基因子結合位點(FBS)His-84氨基酸殘基的刺激下將GTP水解成GDP和Pi;
(5)EF·Tu·GDP對核糖體親和力較低從中釋放;
(6)tRNA進一步發生構象變化,使得其受體臂3‘端和P位點上受體臂3’端距離很近,為肽鏈的轉移作準備;
tRNA的特異性配對大多在此過程中得以篩選確認。如果反密碼子和密碼子不能成功匹配,會影響以下幾個因素導致入位不能完成:
①由於tRNA底部密碼子配對有誤,結構異常,EF·Tu頂部的GTP水解位點不能和HBS完全接觸,導致GTP水解不能發生;
②錯誤的結構,導致tRNA不能發生正確的構象變化,EF·Tu無法釋放;
③由於錯誤的配對,入位時產生的壓力底部無法承受,導致入位失敗。
B. 肽鏈的轉移
肽鏈的轉移化學本質即氨基和羧基的脫水縮合。很長一段時間人們普遍認為核糖體的rRNA保守序列對於脫水縮合起到關鍵的化學催化作用,後事實證明,rRNA僅僅起到結構上的物理催化(上千個序列共同作用而形成的結構),和其任何特定保守序列都沒有完全的直接關係。而tRNA的結構被認為起到了很重要的化學催化作用。
C. 核糖體的移位
(1)肽鏈轉移后,兩個tRNA進行在不穩定的常態和扭轉態(hybrid)之間進行擺動;
(2)EF·G·GTP在tRNA處於扭轉態時結合到A位點上部,鎖定扭轉態結構;
(3)在HBS中His-84殘基的刺激下,GTP發生水解,產生EF·G·GDP·Pi,伴隨著EF·G的構象改變,結構深入A位點,一方面解鎖扭轉態結構,一方面將兩個tRNA推向E位點和P位點;
(4)Pi從EF·G上釋放,使得後者構象再次發生變化,回歸最初的結合位置;
(5)EF·G·GDP對核糖體親和力較弱,從核糖體上釋放;
(6)進入E位點的tRNA不再維持和mRNA的配對,很快得以釋放。
註:扭轉態即tRNA底部依然在該位點,但頂部已經進入下一位點的扭曲狀態。
本過程細胞主要需完成以下目標:
(1)使翻譯停止,不再有新的氨基酸摻入;
(2)釋放合成的多肽鏈;
(3)釋放結合在mRNA上的各組分;
(4)確保核糖體大小亞基以及重要因子的重複利用。
原核細胞和真核細胞在此過程的處理上有明顯不同,下面將分開介紹。
(一)原核細胞
A.肽鏈的釋放
(1)釋放因子RF1/2 (tRNA結構類似)結合A位點,識別並匹配終止密碼子;
(2)RF1/2的GGQ 基序(tRNA受體臂結構類似)催化肽鏈的脫離(以HOH替代HO-進行反應);
(3)RF1/2進一步招募RF3·GDP結合到核糖體大亞基上;
(4)RF3將GDP換成GTP(鳥苷酸交換因子活性),這一過程伴隨的構象改變釋放RF1/2,使其脫離核糖體;
(5)RF3利用自身的GTP水解活性水解GTP回歸GDP結合狀態,隨後從大亞基上脫離。
B. 兩個tRNA的釋放
(1)因子RRF(扭轉態tRNA結構類似)結合到空出來的A位點;
(2)此時具有扭轉態tRNA類似構象的複合物引誘EF·G·GTP結合到大亞基位置並鎖定扭轉態結構;
(3)EF·G·GTP水解GTP轉化為EF·G·GDP·Pi狀態,這一步伴隨構象改變解鎖原本的扭轉態結構,將原本E位點的tRNA推出核糖體,將P位點的tRNA推到E位點,將RRF從A位點推到P位點;
(4)Pi從EF·G釋放,導致構象進一步變化,EF·G脫離,而E位點的tRNA由於較低親和性也從核糖體脫離,RRF由於對P位點的親和性低,也脫離。
C. 大小亞基的分離
(1)由於此時的大小亞基三個位點全空了,翻譯起始因子IF3得以進入結合到E位點,降低大小亞基之間的親和性,使其分離;
(2)IF3繼續結合在小亞基上,準備下一輪起始。
(二)真核細胞
A. 肽鏈的釋放
(1)eRF3充當類似於eEF1(或EF-Tu)的作用,以GTP結合狀態結合到eRF1/2上;
(2)通過eRF3的介導,eRF1/2被運輸到A位點;
(3)eRF1/2識別終止密碼子(類似於tRNA的密碼子配對),正確的構象傳遞使得核糖體FBS和eRF3的GTP結合位點互作;
(4)FBS刺激GTP水解,eRF3轉為GDP結合態;
(5)eRF3·GDP很快從核糖體上脫離,伴隨著eRF1/2的類似於tRNA的入位過程;
(6)eRF1/2的GGQ基序催化肽鏈的脫離;
B. 剩餘組分的脫離
Rli·ATP具有轉位酶活性,通過水解ATP沿著mRNA轉位,以一種類似於轉錄終止的方式,將其他組分從mRNA上推離下來。
目前關於真核細胞翻譯過程有普遍的模型。其認為eIF4G可以結合mRNA的5‘端poly-A尾,進而將mRNA形成閉環,結合多個核糖體,以提高效率。且在此模型下,核糖體在翻譯完畢解離后,可直接在起點重新起始,更符合核糖體的循環利用規律。
任何體內的生物反應都必須在調控的作用下,才有意義。翻譯的調控是十分精密複雜的。在原核生物里翻譯調控的基本單位不是單個的mRNA而是mRNA中的單個閱讀框。以ATP合成酶為例,在原核生物里,該酶包含A、B、C、D、E、F、G、H等多個亞基,其基因拷貝均為一份,在轉錄時轉錄到同一個mRNA上。而實際每個酶中各個亞基的比例並非均等,如E亞基的需求量是C亞基的10倍。這就必須要求在翻譯層面進行調控。
相比於轉錄調控,翻譯調控更迅速、更高效,且可以通過對翻譯的調控進一步控制不同蛋白的空間特異性。同時翻譯調控也可以回復錯誤轉錄造成的危害。
無論針對翻譯、轉錄還是複製,研究調控首先要研究一個生理過程的限速步驟。限速步驟的調控才具有重要的生物學意義。在這裡要引入一個新的單位:核糖體密度(RD)。其計算公式如下:
其中 表示核糖體數量,表示ORF的單位長度。單位是。一般情況下,RD的最大值在1/35左右,而平均RD值在1/154。特別注意的是,只有72個基因表達的轉錄本的RD大於1/50。由於RD本質上反映的是轉錄延伸的速率,其數值之間相差不大表明轉錄延伸並非主要的限速(調控)步驟。由此,翻譯的限速步驟應當是翻譯的起始,而調控也都集中在起始步驟。
(一)原核細胞
在整體上,原核細胞可通過改變核糖體結合位點(RBS)序列或者在RBS鄰域製造二級結構來阻止小亞基和mRNA的結合進而阻止翻譯的起始。一方面由於RBS序列固定,改變其序列將會造成所有mRNA停止翻譯。另一方面由於改變序列並非快速準確的調控方法,針對單個轉錄本,原核細胞傾向於採取以下幾種方式進行調控:
(1)蛋白依賴的——通過蛋白質特異性結合到RBS和起始密碼子AUG之間的序列以通過物理方式阻止小亞基和mRNA結合,但該特異性序列不能包括RBS(否則將可能會導致所有轉錄本停止翻譯)。
(2)mRNA依賴的——通過ORF和下游序列的雜交來阻止該ORF的起始。但如果該ORF上游有其他開放起始位點,則該ORF可以被上游翻譯啟動(核糖體翻譯到下游時,將其解旋),這樣的好處是當且僅當第一個ORF翻譯時,第二個ORF才可以翻譯。
(3)其他——在特殊情況下,原核細胞也可以通過特殊的負反饋途徑和一些特殊的小RNA(sRNA)進行調控。
以核糖體蛋白的翻譯調控為例,我們來討論一個典型的mRNA、蛋白依賴混合的負反饋調控模式。當周圍存在自由rRNA時,新生核糖體蛋白結合rRNA的相應位點來促使其摺疊形成適當結構,此時核糖體蛋白正常翻譯。而當周圍存在多餘的核糖體蛋白時,該蛋白會結合到核糖體蛋白轉錄本的ORF和RBS之間,以促進鈣ORF和下游序列的雜交,抑制翻譯起始。
而其他的調控方法內,最為典型的是核糖開關。核糖開關是一類小mRNA,其能翻譯出一些蛋白質,後者催化產生一些類別的SAM(小活性分子)。當SAM存在時,可促使mRNA發生構象變化,在ORF和RBS等關鍵位點處形成二級結構,阻遏翻譯起始。當SAM不存在時,阻遏不能形成,翻譯得以繼續。
一些小RNA(sRNA)也可以起到翻譯調控的作用。原核生物的小RNA大小基本在80-110核苷酸。它可以通過自身和RBS互補的序列來解除RBS參與的自體雜交,進而回復翻譯。它也可以直接和開放的RBS雜交阻遏翻譯,具有典型的兩重性。
(二)真核細胞
值得注意的是,雖然在原核生物細胞內,翻譯的起始過程依然有IF1、IF2、IF3三類因子的參與(真正耗能的步驟是IF2介導的起始tRNA入位和大亞基招募),但原核細胞幾乎沒有以這些蛋白因子為靶點進行的調控模式。在真核細胞內,由於大量翻譯起始因子的參與,大量對於翻譯的調控也是以這些蛋白因子為靶點進行的。
和原核細胞類似,真核細胞的翻譯調控也可分為基因特異性調控和全局調控。其靶點主要有三個:起始tRNA的入位、mRNA-eIF4複合物的形成、mRNA的掃描。
最常見的全局調控之一是激酶(如GCN2)磷酸化eIF2,使原本使eIF2·GDP轉為eIF2·GTP的eIF2B失去鳥苷酸交換因子活性,反應不能發生,eIF2·GTP減少,tRNA入位減少,細胞內全局翻譯速率降低。
另一種常見的全局調控是利用4EBP(eIF4E結合蛋白)與eIF4G競爭,識別並結合eIF4E。而激酶可以磷酸化4EBP以使其不具有結合4E的活性。這一通路常被視作一些營養通路(如mTOR)的效應下游。
基因特異性調控主要依賴於mRNA的特有序列。常見的例子是Bruno蛋白可以特異性結合一些mRNA下游元件,而與此同時Cup蛋白與eIF4E結合。由於Bruno可以和Cup互作,進而形成mRNA閉合環路,阻止翻譯的進一步發生。
另一個特異性調控的例子是鐵蛋白的翻譯調控。由於鐵蛋白主要參與二價鐵離子的代謝,在二價鐵離子存在時需要被高度表達以應激,防止其進一步破壞細胞。鐵蛋白mRNA是一類帶有IRE序列的mRNA。當鐵離子不存在時,IRP(IRE結合蛋白)可結合IRE序列,阻礙4A因子對mRNA的AUG的掃描。而當鐵離子存在時,IRP被釋放,掃描得以回復。
約50%的智人(Homo Sapiens)基因帶有上游起始密碼子(上游AUG)。儘管真核細胞每條mRNA只表達一個蛋白,但其依然可以帶有多個上游可讀框和上游AUG,只是其並不翻譯產生具有生物學意義的蛋白。理論上,真核細胞會翻譯其mRNA上游到下游掃描遇到的第一個AUG,並且在翻譯完成後解離。這意味著上游AUG的存在將阻礙下游AUG的啟動,進而阻礙具有生物學意義的蛋白的表達。但這並不是絕對的,上游AUG/ORF的確起到負調控的作用,但不能完全阻止下游ORF的翻譯。其具體機制尚不完全清楚。
上游密碼子的克服
一般認為核糖體可以通過以下兩種方式來克服上游AUG的阻礙:
(1)直接略過第一個AUG。
一般有兩種猜想支持這一個方法:Kozak序列和二級結構。由於一部分上游AUG帶有較為保守的Kozak序列:A/GNNAUGG,或許生物可以通過某種機制識別這一序列進而直接略過上游AUG的翻譯。同時,也有研究表明在上游AUG周圍出現二級結構可以使得其被核糖體略過而不啟動翻譯,而其在機制上並不容易理解,因為理論上eIF4A會解旋所有二級結構,保證翻譯不受影響。
(2)翻譯在上游ORF終止后重新啟動。
由於ORF大多很短(只有3-10個氨基酸),核糖體小亞基或許在翻譯結束後繼續和mRNA互作,並持續掃描而並不解離下來。一般認為此類機制下的翻譯重啟也會受到下游終止密碼子的影響。
上游密碼子的調控作用
已有證據表明,上游密碼子是細胞內經常採取的一種通用調節翻譯的機制。下面以GCN4在響應低氨基酸應激過程中的翻譯調控為例進行討論。
GCN4作為一類蛋白因子,可以上調一大類氨基酸合成酶的表達。GCN4的mRNA具有非常特殊的5'端結構,大體如下:
GCN4的mRNA具有非常特殊的5'端結構
突變4個uORF的起始密碼子AUG,依次建立了4個突變組M1~4,同時再另取一組將其ORF2~4起始密碼子AUG均突變,建立了第5個突變組M5,保留一組野生型(WT)作對照,實驗觀察GCN4在高氨基酸和低氨基酸下的表達情況,可以得到如下結果(+表示表達,-表示不表達):
| 低氨基酸 | 高氨基酸 | |
| M1 | - | - |
| M2 | + | - |
| M3 | + | - |
| M4 | + | +/- |
| M5 | + | + |
| WT | + | - |
其調控機制是這樣的:
GCN2是GCN4的同家族蛋白,可以響應低氨基酸狀態,進一步作為激酶磷酸化eIF2。磷酸化的eIF2可以抑制eIF2B的鳥苷酸交換因子活性。不同尋常的是,核糖體在翻譯經過ORF1起始密碼子后,部分翻譯起始因子得以保留,待經過終止密碼子后,大亞基解離,小亞基和部分翻譯起始因子繼續保留在mRNA上。
①當氨基酸含量充足時,激酶GCN2 不響應,eIF2 未被磷酸化,eIF2B 鳥苷酸交換因子活性正常,將 eIF2 由 GDP 結合態轉變為 GTP 結合態,後者狀態的 eIF2 正常結合氨醯-tRNA 並轉運至核糖體 P 位點;由於核糖體度過 ORF1 后可繼續結合併在 mRNA 上移位。由於 eIF2·GTP·tRNA 三元複合物較充足,核糖體容易在 uORF 區域結合 tRNA 進而對 ORF2~4 中的任意一個進行翻譯產生不具有生物學意義的短肽,而翻譯后的核糖體從 mRNA 上脫離,不接觸 dORF,GCN4 不表達;
②當氨基酸含量不足時,激酶GCN2 應激進而具有激酶活性,磷酸化 eIF2,磷酸化的 eIF2 無法被 eIF2B 交 換磷酸鳥苷,仍然處於 GDP 結合態,GTP 結合態的 eIF2 減少,則 eIF2·GTP·tRNA 三元複合物數量減少;此時在 mRNA 上移位的核糖體小亞基不易遇到合適的 tRNA 並開始翻譯,有更大機會度過 uORF 區域進入 大間隔區;由於大間隔區長度大,賦予了核糖體小亞基更多的時間讓其有更大機會遇到合適的 tRNA 並在 dORF 區域開始翻譯,GCN4 進而表達。
翻譯過程需要的原料:mRNA、tRNA、20種氨基酸、能量、酶、核糖體。
翻譯的過程大致可分作三個階段:起始、延長、終止。翻譯主要在細胞質內的核糖體中進行,氨基酸分子在氨基醯-tRNA合成酶的催化作用下與特定的轉運RNA結合併被帶到核糖體上。生成的多肽鏈(即氨基酸鏈)需要通過正確摺疊形成蛋白質,許多蛋白質在翻譯結束后還需要在內質網上進行翻譯后修飾才能具有真正的生物學活性。
遊離的鹼基以mRNA為直接模板,tRNA為氨基酸運載體,核蛋白體為裝配場所,共同協調完成蛋白質生物合成的過程。在翻譯過程中mRNA上的每個三聯體密碼子對應tRNA上的一個三聯體反密碼子,且這個反密碼子只對應一個氨基酸,但是一個氨基酸可有多組密碼子(密碼子具有簡併性)來表示。
一個激活的tRNA進入核糖體的A位(A位用於接受氨基酸,可記憶為Accept)與mRNA相配,肽醯轉移酶在鄰近的氨基酸間建立一個肽鍵,此後在P位(peptidyl-tRNA site,即肽醯基位點,也叫做“供點”)上的氨基酸離開它的tRNA與A位上的tRNA結合,核糖體則相對於mRNA向前滑動,原來在A位上的tRNA移動到P位上,原來在P位上的空的tRNA移動到E位(釋放位點,記憶為Emission)上,然後在下一個tRNA進入A位之前被釋放。
當核糖體沿著mRNA分子移動到A位出現終止密碼子時,沒有相應的氨醯tRNA與之對應,一種釋放因子(Release Factor,RF)與終止密碼子及核糖體A位結合,另一種釋放因子隨之結合,改變肽醯轉移酶的特異性,催化P位肽醯tRNA水解,從而使肽鏈從核糖體上釋放。
翻譯過程嚴格按照鹼基互補配對原則進行。值得注意的是,轉錄組成RNA的鹼基中,沒有T(胸腺嘧啶),而是U(尿嘧啶),所以配對原則如下所示。
DNA -> RNA
A ,腺嘌呤-> U,尿嘧啶
T ,胸腺嘧啶-> A ,腺嘌呤
G ,鳥嘌呤-> C,胞嘧啶
C,胞嘧啶-> G,鳥嘌呤.
由於密碼子的擺動性,反密碼子第1位鹼基與密碼子第3三位鹼基之間配對稱多樣性,可能配對方式如下:
I ->A,U,C
U ->A,G
G ->C,U
其中,I代表次黃嘌呤核苷(inosine)。
(Commaless)
編碼蛋白質氨基酸序列的各個三聯體密碼連續閱讀,密碼間既無間斷也無交叉。
(degeneracy)
遺傳密碼中,除色氨酸和甲硫氨酸僅有一個密碼子外,其餘氨基酸有2、3、4個或多至6個三聯體為其編碼。
(universal)
蛋白質生物合成的整套密碼,從原核生物到人類都通用。
已發現少數例外,如動物細胞的線粒體、植物細胞的葉綠體。
密碼的通用性進一步證明各種生物進化自同一祖先。
(wobble)
轉運氨基酸的tRNA的反密碼需要通過鹼基互補與mRNA上的遺傳密碼反向配對結合,但反密碼與密碼間不嚴格遵守常見的鹼基配對規律,稱為擺動配對。
(direction)
起始密碼總位於編碼區5′末端,而終止密碼位於3′末端,每個密碼的三個核苷酸也是5′至3′方向閱讀,不能倒讀。

基本信息
- 中文名
- 翻譯
- 外文名
- translation
- 所屬學科
- 生物學
- 根據
- 鹼基的排列順序
- 作用
- 生成氨基酸序列