核糖核酸
遺傳信息載體
核糖核酸(英語:Ribonucleic acid,縮寫:RNA)存在於生物細胞以及部分病毒、類病毒中的遺傳信息載體。
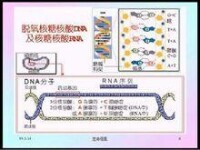
核糖核酸
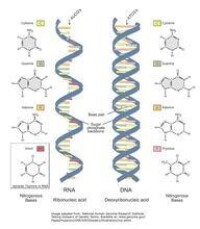
RNA由核糖核苷酸經磷酯鍵縮合而成長鏈狀分子。一個核糖核苷酸分子由磷酸,核糖和鹼基構成。RNA的鹼基主要有4種,即A腺嘌呤,G鳥嘌呤,C胞嘧啶,U尿嘧啶。其中,U尿嘧啶取代了DNA中的T胸腺嘧啶而成為RNA的特徵鹼基。
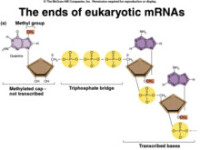
mRNA的功能就是把DNA上的遺傳信息精確無誤地轉錄下來,然後再由mRNA的鹼基順序決定蛋白質的氨基酸順序,完成基因表過程中的遺傳信息傳遞過程。在真核生物中,轉錄形成的前體RNA中含有大量非編碼序列,大約只有25%序列經加工成為mRNA,最後翻譯為蛋白質。因為這種未經加工的前體mRNA(pre-mRNA)在分子大小上差別很大,所以通常稱為不均一核RNA(heterogeneousnuclearRNA,hnRNA)。
如果說mRNA是合成蛋白質的藍圖,則核糖體是合成蛋白質的工廠。但是,合成蛋白質的原材料——20種氨基酸與mRNA的鹼基之間缺乏特殊的親和力。因此,必須用一種特殊的RNA——轉移RNA(transferRNA,tRNA)把氨基酸搬運到核糖體上,tRNA能根據mRNA的遺傳密碼依次準確地將它攜帶的氨基酸連結起來形成多肽鏈。每種氨基酸可與1-4種tRNA相結合,已知的tRNA的種類在40種以上。
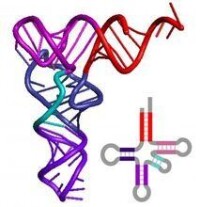
tRNA
1969年以來,研究了來自各種不同生物,如:如酵母、大腸桿菌、小麥、鼠等十幾種tRNA的結構,證明它們的鹼基序列都能摺疊成三葉草形二級結構,而且都具有如下的共性:
①5’末端具有G(大部分)或C。
②3’末端都以ACC的順序終結。
③有一個富有鳥嘌呤的環。
⑤有一個胸腺嘧啶環。
核糖體RNA(ribosomalRNA,rRNA)是組成核糖體的主要成分。核糖體是合成蛋白質的工廠。在大腸桿菌中,rRNA量占細胞總RNA量的75%-85%,而tRNA佔15%,mRNA僅佔3-5%。
rRNA
S為沉降係數(sedimentationcoefficient),當用超速離心測定一個粒子的沉澱速度時,此速度與粒子的大小直徑成比例。5S含有120個核苷酸,16S含有1540個核苷酸,而23S含有2900個核苷酸。而真核生物有4種rRNA,它們分子大小分別是5S、5.8S、18S和28S,分別具有大約120、160、1900和4700個核苷酸。rRNA是單鏈,它包含不等量的A與U、G與C,但是有廣泛的雙鏈區域。在雙鏈區,鹼基因氫鍵相連,表現為髮夾式螺旋。
rRNA在蛋白質合成中的功能尚未完全明了。但16S的rRNA3’端有一段核苷酸序列與mRNA的前導序列是互補的,這可能有助於mRNA與核糖體的結合。
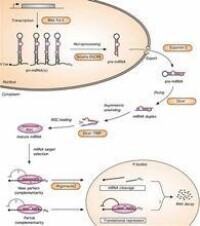
miRNA
(small RNA)
存在於真核生物細胞核和細胞質中,它們的長度為100到300個鹼基(酵母中最長的約1000個鹼基)。多的每個細胞中可含有105~106個這種RNA分子,少的則不可直接檢測到,它們由RNA聚合酶Ⅱ或RNA聚合酶Ⅲ所合成,其中某些像mRNA一樣可被加帽。
主要有兩種類型的小分子RNA:一類是snRNA(small nuclear RNA),存在於細胞核中;另一類是scRNA(small cytoplasmic RNA),存在於細胞質中。
小分子RNA通常與蛋白質組成複合物,在細胞的生命活動中起重要的作用。
①snRNA:
snRNA(smallnuclearRNA,小核RNA)。它是真核生物轉錄后加工過程中RNA剪接體(spilceosome)的主要成分。發現有五種snRNA,其長度在哺乳動物中約為100-215個核苷酸。snRNA一直存在於細胞核中,與40種左右的核內蛋白質共同組成RNA剪接體,在RNA轉錄后加工中起重要作用。某些snRNPs和剪接作用密切相關,它們分別與供體和受體剪接位點以及分支順序相互補。
其中位於核仁內的snRNA稱為核小體RNA(small uncleolar RNA),參與rRNA前體的加工及核糖體亞基的組裝。
②scRNA:
scRNA(small cytoplasmic RNA,細胞質小RNA)主要位於細胞質內,種類較多,參與蛋白質的合成和運輸。SRP顆粒就是一種由一個7SRNA和六種蛋白質組成的核糖核蛋白體顆粒,主要功能是識別信號肽,並將核糖體引導到內質網。
端體酶RNA
反義RNA(antisenseRNA),它參與基因表達的調控。
上述各種RNA分子均為轉錄的產物,mRNA最後翻譯為蛋白質,而rRNA、tRNA及snRNA等並不攜帶翻譯為蛋白質的信息,其終產物就是RNA。
另外還有一種特別的RNA(其分類與上述RNA分類無關)——核酶
核酶(ribozyme)一詞用於描述具有催化活性的RNA,即化學本質是核糖核酸(RNA),卻具有酶的催化功能。核酶的作用底物可以是不同的分子,有些作用底物就是同一RNA分子中的某些部位。核酶的功能很多,有的能夠切割RNA,有的能夠切割DNA,有些還具有RNA連接酶、磷酸酶等活性。與蛋白質酶相比,核酶的催化效率較低,是一種較為原始的催化酶。
大多數核酶通過催化轉磷酸酯和磷酸二酯鍵水解反應參與RNA自身剪切、加工過程,也具有特異性,甚至具有Km值。
其發現是科學家大腸桿菌RNaseP蛋白在切去部分后,在體外高濃度鎂離子的情況下,留下的RNA部分(MIRNA)具有酶活性。
【新型生命暗物質】非編碼RNA(核糖核酸),被稱為生命體中“暗物質”。日前,中國科學技術大學單革教授實驗室發現一類新型環狀非編碼RNA,並揭示了此類非編碼RNA的功能和功能機理。成果發表在國際知名雜誌《自然·結構和分子生物學》上。非編碼RNA是一大類不編碼蛋白質,但在細胞中起著調控作用的RNA分子
。
正如宇宙間存在著許多既看不到也感覺不到的“暗物質”“暗能量”一樣,在生命體這個“小宇宙”中,也存在這樣的神秘“暗物質”—非編碼RNA。
越來越多的證據表明,一系列重大疾病的發生髮展與非編碼RNA調控失衡相關。
環形RNA分子最近數年才引起研究人員注意,而此前的研究主要集中於線性RNA分子。單革教授實驗室發現的新型環狀非編碼RNA,被命名為外顯子-內含子環形RNA。在論文中,他們還對這類新型環狀非編碼RNA為何會成為環形而不是線形分子進行了研究,發現成環序列兩端經常會有互補的重複序列存在。
。
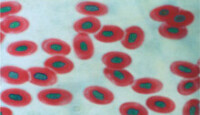
蟾蜍血塗片
由於DNA和RNA在化學組成與分子結構上存在一定的差別,因而對不同的染料有著不同的反應。所以,可以根據這一反應差異來研究細胞中DNA與RNA的分佈情況。
DNA和RNA兩種核酸分子都是多聚體,但是它們的聚合程度有所不同。DNA聚合程度高,易於甲基綠結合;RNA聚合程度低易與吡羅紅結合。所以當吡羅紅與甲基綠混在一起作為染料時吡羅紅與核仁、細胞質中的RNA選擇性結合,從而顯示紅色;甲基綠與染色質中的DNA選擇性結合,從而顯示綠色。綜上所述,RNA對吡羅紅的親和力大,被染成紅色;DNA對甲基綠的親和力大,被染成綠色。
與DNA不同,RNA一般為單鏈長分子,不形成雙螺旋結構,核糖核酸但是很多RNA也需要通過鹼基配對原則形成一定的二級結構乃至三級結構來行使生物學功能。RNA的鹼基配對規則基本和DNA相同,不過除了A-U、G-C配對外,G-U也可以配對。
在細胞中,根據結構功能的不同,RNA主要分三類,即tRNA(轉運RNA),rRNA(核糖體RNA),mRNA(信使RNA)。mRNA是合成蛋白質的模板,內容按照細胞核中的DNA所轉錄;tRNA是mRNA上鹼基序列(即遺傳密碼子)的識別者和氨基酸的轉運者;rRNA是組成核糖體的組分,是蛋白質合成的工作場所。
在病毒方面,很多病毒只以RNA作為其唯一的遺傳信息載體(有別於細胞生物普遍用雙鏈DNA作載體)。
核糖核酸
20世紀90年代以來,又發現了RNAi(RNAinterference,RNA干擾)等等現象,證明RNA在基因表達調控中起到重要作用。
在RNA病毒中,RNA是遺傳物質,植物病毒總是含RNA。近些年在植物中陸續發現一些比病毒還小得多的浸染性致病因子,叫做類病毒。類病毒是不含蛋白質的閉環單鏈RNA分子,此外,真核細胞中還有兩類RNA,即不均一核RNA(hnRNA)和小核RNA(snRNA)。hnRNA是mRNA的前體;snRNA參與hnRNA的剪接(一種加工過程)。自1965年酵母丙氨酸tRNA的鹼基序列確定以後,RNA序列測定方法不斷得到改進。除多種tRNA、5SrRNA、5.8SrRNA等較小的RNA外,尚有一些病毒RNA、mRNA及較大RNA的一級結構測定已完成,如噬菌體MS2RNA含3569個核苷酸。
核糖核酸
類似的謎團,直到美國科學家安德魯·法爾和克雷格·梅洛發現核糖核酸
RNA(核糖核酸)干擾機制才得到科學的解釋。兩位科學家也正是因為1998年做出的這一發現而榮獲2006年的諾貝爾生理學或醫學獎。
上世紀八十年代,托馬斯.R.切赫博士在研究RNA的成熟體結構中,發現了可以自我拼接的RNA催化作用(核糖核苷酸酶),並依此榮獲1989年諾貝爾化學獎。經過多年的深度研究,切赫博士在DNA基因遺傳過程中,發現了有趣的mRNA(信使RNA)和tRNA(轉運RNA),從而揭開了遺傳基因導致出生缺陷、大腦發育、營養吸收、細胞變異以及健康長壽等一系列人類生命密碼的神秘面紗。
mRNA(信使RNA)人類的遺傳信息主要貯存於DNA的鹼基序列中,不過DNA並不直接決定蛋白質的合成。而在真核細胞中,DNA主要貯存於細胞核中的染色體上,而蛋白質的合成場所存在於細胞質中的核糖體上,因此需要有一種中介物質,才能把DNA上控制蛋白質合成的遺傳信息傳遞給核糖體。切赫博士把這種起著傳遞遺傳信息作用的特殊RNA。稱為信使RNA(messenger RNA,mRNA)。
簡單的說,mRNA就是為了完成基因表達過程中的遺傳信息傳遞。
令人遺憾的是,在遺傳轉錄形成的過程中,僅有25%序列經加工成為mRNA,其餘的均呈現非編碼序列的前體mRNA形式,這些形式的mRNA在分子大小上差別很大,是導致出生缺陷、大腦發育、營養吸收、細胞變異以及健康長壽等一系列問題的基因遺傳因素的關鍵所在。
切赫博士歷經20年升華鑽研,成果破譯了mRNA編碼序列信息奧秘,通過特殊的生物干預手段,優化mRNA的序列加工,篩查和剔除基因排列誘發基因和細胞突變的序列,不僅確保mRNA的序列加工的有效與增強,而且從根本上避免不良基因傳遞或傳遞序列問題引發細胞突變等一系列遺傳問題的發生。
mRNA編碼序列信息的成果破譯,奠定了OMG配方鹽技術的可行性基礎。
法爾和梅洛的發現
科學家在矮牽牛花實驗中所觀察到的奇怪現象,其實是因為生物體內某種特定基因“沉默”了。導致基因“沉默”的機制就是RNA干擾機制。
此前,RNA分子只是被當作從DNA(脫氧核糖核酸)到蛋白質的“中間人”、將遺傳信息從“藍圖”傳到“工人”手中的“信使”。但法爾和梅洛的研究讓人們認識到,RNA作用不可小視,它可以使特定基因開啟、關閉、更活躍或更不活躍,從而影響生物的體型和發育等。
諾貝爾獎評審委員會在評價法爾和梅洛的研究成果時說:“他們的發現能解釋許多令人困惑、相互矛盾的實驗觀察結果,並揭示了控制遺傳信息流動的自然機制。這開啟了一個新的研究領域。”
siRNA的作用原理
核糖核酸
RNA干擾技術的前景
RNA干擾技術不僅是研究基因功能的一種強大工具,不久的未來,這種技術也許能用來直接從源頭上讓致病基因“沉默”,以治療癌症甚至艾滋病,在農業上也將大有可為。從這個角度來說,“沉默”真的是金。美國哈佛醫學院研究人員已用動物實驗表明,利用RNA干擾技術可治癒實驗鼠的肝炎。
儘管尚有一些難題阻礙著RNA干擾技術的發展,但科學界普遍對這一新興的生物工程技術寄予厚望。這也是諾貝爾獎評審委員會為什麼不堅持研究成果要經過數十年實踐驗證的“慣例”,而破格為法爾和梅洛頒獎的原因之一。
諾貝爾生理學或醫學獎評審委員會主席戈蘭·漢松說:“我們為一種基本機制的發現頒獎。這種機制已被全世界的科學家證明是正確的,是給它發個諾貝爾獎的時候了。”
轉錄是指DNA的雙鏈解開,使RNA聚合酶可依照DNA上的鹼基序列合成相對應之信使RNA(mRNA)的過程.在人體需要酵素或是蛋白質時,都會需要進行此過程,才能藉由信使mRNA,將密碼子帶出核模外.好讓核糖體進一步的利用信使RNA(mRNA)來翻譯,合成所需之蛋白質‧DNA的鹼基有A(腺嘌呤)、G(鳥嘌呤)、C(胞嘧啶)、T(胸腺嘧啶),而RNA之鹼基無T(胸腺嘧啶),取而代之的是U(尿嘧啶),也就是有A(腺嘌呤)、G(鳥嘌呤)、C(胞嘧啶)、U(尿嘧啶).在DNA中,A與T以兩條氫鍵連結,G與C以三條氫鍵連結,但RNA只有U而無T,所以在轉錄時DNA上的若是A,mRNA就會是U,也就是取代原本T的位置‧如下圖所示,右邊DNA的一股鹼基序列若為‘AAACCG’,而左方的DNA因配對而就會成‘TTTGGC’,但因RNA無T這個鹼基,只有U,因此合成出來的mRNA對應之序列就為‘UUUGGC’因為DNA太大,無法出入核膜(細胞核的膜),所以才需要有mRNA的出現,讓mRNA可穿過核孔(核膜上的孔洞)到達細胞質進行翻譯(核糖體合成蛋白質的過程),因此,轉錄對不管是人類還是動物甚至是細菌都是不可或缺的重要反應。
遊離在細胞質中的各種氨基酸,就以mRNA為模版合成具有一定氨基酸順序的蛋白質,這一過程叫翻譯。
首先氨基酸與tRNA結合生成氨醯-tRNA
然後是多肽鏈的起始:mRNA從核到胞質,在起始因子和Mg的作用下,小亞基與mRNA的起始部位結合,甲硫氨醯(蛋氨酸)—tRNA的反密碼子,識別mRNA上的起始密碼AuG(mRNA)互補結合,接著大亞基也結合上去,核糖體上一次可容納二個密碼子。(原核生物中為甲醯甲硫氨醯)
再是多肽鏈的延長:第二個密碼對應的氨醯基—tRNA進入核糖體的A位,也稱受位,密碼與反密碼的氫鍵,互補結合。在大亞基上的多肽鏈轉移酶(轉肽酶)作用下,供位(P位)的tRNA攜帶的氨基酸轉移到A位的氨基酸后並與之形成肽鍵(—CO-NH—),tRNA脫離P位並離開P位,重新進入胞質,同時,核糖體沿mRNA往前移動,新的密碼又處於核糖體的A位,與之對應的新氨基醯-tRNA又入A位,轉肽鍵把二肽掛於此氨基酸后形成三肽,ribosome又往前移動,由此漸進漸進,如此反覆循環,就使mRNA上的核苷酸順序轉變為氨基酸的排列順序。
最後是多肽鏈的終止與釋放:肽鏈的延長不是無限止的。當mRNA上出現終止密碼時(UGA、U氨基酸和UGA),就無對應的氨基酸運入核糖體,肽鏈的合成停止,而被終止因子識別,進入A位,抑制轉肽酶作用,使多肽鏈與tRNA之間水解脫下,順著大亞基中央管全部釋放出,離開核糖體。同時大小亞基與mRNA分離,可再與mRNA起始密碼處結合,也可遊離於胞質中或被降解,mRNA也可被降解。
2013年5月新一期《科學》雜誌上報告說,英國約翰·英尼斯中心的研究人員發現的這種核糖核酸名為COOLAIR,是一種反義長鏈非編碼核糖核酸。研究人員以模式植物擬南芥作為研究對象,通過遺傳篩選和基因克隆等手段,發現COOLAIR受到一種叫做R環的特殊結構的影響。R環是由一條脫氧核糖核酸(DNA)與核糖核酸雜合鏈以及一條單鏈DNA所形成的特殊基因組結構,一般在基因表達轉錄核糖核酸時可以形成瞬時的R環,但很快會被去除。而他們觀察發現,R環能夠通過抑制COOLAIR發揮作用,從而讓擬南芥提前開花。
2021年5月,中科院發現基因魔剪新系統,被廣泛地應用於RNA敲低、RNA單鹼基編輯、RNA定點修飾、RNA活細胞示蹤以及核酸檢測領域。
mRNA
mRNA含A、U、G、C四種核苷酸,每三個相聯而成一個三聯體,即密碼,代表一個氨基酸的信息,故按數學中排列組合法則計算,可形成43=64個不同的密碼。
64個密碼中,61個密碼分別代表各種氨基酸。每種氨基酸少的只有一個密碼,多的可有6個,但以2個及4個的居多數。此外,UAA、UAG、UGA這三個密碼是肽鏈合成的終止信號,不代表任何氨基酸。在真核生物中,AUG既是甲硫氨酸的密碼,又是肽鏈合成的起始信號;而在原核生物中,GUG(在真核生物中是纈氨酸的密碼)和AUG樣,都是甲醯甲硫氨酸的密碼和肽鏈合成的起始相號。可見,除GUG外,所有的密碼從細菌到高等生物都能適用,這一點為生物的共同起源學說提供了有力的佐證。
必須指出:⑨在mRNA整個分子中,從起始信號直至終止信號,其密碼的三聯體是連續的,密碼與密碼之間沒有間隔的核苷酸;②起始信號AUG並非是mRNA的起始(5′端),而可以和5′端間隔若干個核苷酸;而且終止信號也不在mRNA的3′端。
tRNA
作為“搬運工具”的tRNA有很多種,體內20種氨基酸都有其自已特有的tRNA,所以,tRNA的種類不少於20種。tRNA在ATP供應能量和酶的作用下,可分別與特定的氨基酸結合。每個tRNA都有一個由三個核苷酸編成的“反密碼”。這個反密碼可以根據鹼基配對的原則與mRNA上對應的密碼配對,而且只有當反密碼與mRNA上的密碼相對應時才能配合,否則就“格格不入”。所以在翻譯時,帶著不同氨基酸的各個tRNA就能準確地在mRNA分子上“對號入座”,依次與典密碼相合,這就保證了氨基酸能排列成一定的順序。
tRNA上的反密碼當然應能識別mRNA上相應的、互補的密碼,並與之配對。然而用提純的tRNA來進行實驗時,發現一種tRNA能夠識別幾種密碼。例如,丙氨酸tRNA,其反密碼為IGC(5′>3′),可以識別三種密碼。
rRNA
rRNA與多種蛋白質分子共同構成核蛋白體。核蛋白體相當於“裝配機”,能促使tRNA所攜帶的氨基醯基縮合成肽。核蛋白體附著在mRNA上,並沿著mRNA長鏈的起始信號向終止信號移動。至於rRNA在蛋白質生物合成中的具體作用還不清楚。
基本信息
- 中文名
- 核糖核酸
- 外文名
- Ribonucleic Acid
- 別名
- RNA
- 本質
- 長鏈狀分子
- 鹼基
- A、G、C、U
- 原則
- 鹼基互補配對原則
- 過程
- 轉錄、翻譯等
- 構成
- 磷酸、核糖和鹼基
- 作用
- 引導蛋白質合成