碳四植物
CO2同化的最初產物
CO2同化的最初產物不是光合碳循環中的三碳化合物3-磷酸甘油酸,而是四碳化合物蘋果酸或天門冬氨酸的植物。又稱C4植物。如玉米、甘蔗、高粱、莧菜等。而最初產物是3-磷酸甘油酸的植物則稱為碳三植物(C3植物)。
碳四植物常寫作C4植物。生長過程中從空氣中吸收二氧化碳首先合成蘋果酸或天門冬氨酸等含四個碳原子化合物的植物,如玉米、甘蔗等。而小麥、水稻等作物先合成磷甘油酸等三碳原子分子,為C3植物。C4植物較之C3植物具有生長能力強、二氧化碳利用率高、需水分量少等許多優點。禾本科經濟植物中約有300種屬C4植物。用雜交法或細胞融合法培育雜交水稻等,使其具有跟C4植物相近的二氧化碳攝取能力的育種工作一直在進行。培育近似C4植物的新品種,對全世界糧食狀況的改善具有重大意義。
一般植物中,二氧化碳同化時固定的第一個產物是具有3個碳原子的磷酸甘油酸,採用這種途徑的植物稱碳3植物,,如大豆、棉花、小麥和稻等。而有些植物中,二氧化碳固定的第一個產物是具有4個碳原子的雙羧酸,採用這種途徑的植物稱碳4植物,,如玉米、高粱和甘蔗等。二氧化碳首先在葉肉細胞內被固定在四碳雙羧酸中,然後被運輸到維管束鞘細胞中脫羧,放出的二氧化碳被Rubisco催化的羧化反應再次固定,該途徑實際上是在二氧化碳同化的基礎上增加了一個四碳雙羧酸循環,這個循環像一個二氧化碳泵,使Rubisco羧化部位的二氧化碳濃度比碳3植物的高很多,從而減少光呼吸,因而碳4植物在強光下具有比碳3植物更高的光合效率。
有些耐乾旱的植物如景天、仙人掌等的光合碳同化過程與碳4植物類似,先將二氧化碳固定並還原成四碳雙羧酸,但它們不在兩類細胞間運輸,而是將兩次二氧化碳固定的時間錯開:夜間氣孔開放,吸收的二氧化碳固定於四碳雙羧酸中;白天氣孔關閉,四碳雙羧酸脫羧釋放的二氧化碳再次固定在磷酸甘油酸中,並同化成磷酸丙糖,避免了白天強烈的水分蒸騰,更有利於植物在乾旱環境中生存。
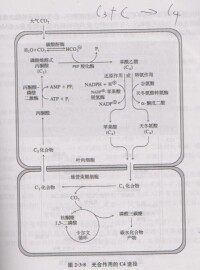
光合作用的C4途徑
葉肉細胞里的磷酸烯醇式丙酮酸(PEP)經PEP羧化酶的作用,與CO2結合,形成蘋果酸或天門冬氨酸。這些四碳雙羧酸轉移到鞘細胞里,通過脫羧酶的作用釋放CO2,後者在鞘細胞葉綠體內經核酮糖二磷酸(RuBP)羧化酶作用,進入光合碳循環。這種由PEP形成四碳雙羧酸,然後又脫羧釋放CO2的代謝途徑稱為四碳途徑。其葉肉細胞中,含有獨特的酶,即磷酸烯醇式丙酮酸碳氧化酶,使得二氧化碳先被一種三碳化合物--磷酸烯醇式丙酮酸同化,形成四碳化合物草醯乙酸鹽,這也是該暗反應類型名稱的由來。這草醯乙酸鹽在轉變為蘋果酸鹽后,進入維管束鞘,就會分解釋放二氧化碳和一分子丙酮酸。二氧化碳進入卡爾文循環,后同C3進程。而丙酮酸則會被再次合成磷酸烯醇式丙酮酸,此過程消耗ATP。
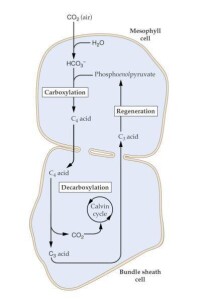
碳四植物
已經發現的四碳植物約有2000種,廣泛分佈在植物的20個不同的科中。它們大都起源於熱帶。因為四碳植物能利用強日光下產生的ATP推動PEP與CO2的結合,提高強光、高溫下的光合速率,在乾旱時可以部分地收縮氣孔孔徑,減少蒸騰失水,而光合速率降低的程度就相對較小,從而提高了水分在四碳植物中的利用率。這些特性在乾熱地區有明顯的選擇上的優勢。
C4植物與C3植物的一個重要區別是C4植物的CO2補償點很低,而C3植物的補償點很高,所以C4植物在CO2含量低的情況下存活率更高。
C4植物主要是那些生活在乾旱熱帶地區的植物。在這種環境中,植物若長時間開放氣孔吸收二氧化碳,會導致水分通過蒸騰作用過快的流失。所以,植物只能短時間開放氣孔,二氧化碳的攝入量必然少。植物必須利用這少量的二氧化碳進行光合作用,合成自身生長所需的物質。
在C4植物葉片維管束的周圍,有維管束鞘圍繞,這些維管束鞘細胞里有葉綠體,但裡面並無基粒或基粒發育不良。在這裡,主要進行卡爾文循環。
該類型的優點是,二氧化碳固定效率比C3高很多,有利於植物在乾旱環境生長。C3植物行光合作用所得的澱粉會貯存在葉肉細胞中,因為這是卡爾文循環的場所,而維管束鞘細胞則不含葉綠體。而C4植物的澱粉將會貯存於維管束鞘細胞內,因為C4植物的卡爾文循環是在此發生的。
碳四植物
這種能夠固定CO2的酶,叫做磷酸烯醇式丙酮酸羧化酶,簡稱PEP羧化酶。
釋放出來的CO2先被一個C5固定,然後很快形成兩個C3。在有關酶的催化作用下,一些C3接受ATP和NADPH釋放出的能量並且被NADPH還原,然後經過一系列複雜的變化,形成糖類等有機物;另一些C3則經過複雜的變化,又形成C5,從而使暗反應階段的化學反應不斷地進行下去。C4釋放出的CO2的變化情況,與C3植物暗反應階段的變化情況相同。丙酮酸則再次進入到葉肉細胞中的葉綠體內,在有關酶的催化作用下,通過ATP提供的能量,轉化成PEP,PEP則可以繼續固定CO2(如圖)。
由此可見,C4植物的光合作用中既有C4途徑,又有C3途徑,前者發生在葉肉細胞的葉綠體內,後者發生在維管束鞘細胞的葉綠體內,兩者共同完成二氧化碳的固定。
同C3途徑中有關的酶與CO2的親和力相比,C4途中PEP羧化酶與CO2的親和力約高60倍。
C4植物利用PEP將CO2固定在C4中,C4經過一系列的變化后,又把CO2釋放出來,這有什麼意義呢?原來,C4途徑中能夠固定CO2的那種酶,對CO2具有很強的親合力,可以促使PEP把大氣中濃度很低的CO2固定下來,並且使C4集中到維管束鞘細胞內的葉綠體中,供維管束鞘細胞內葉綠體中的C3途徑利用。科學家們把C4植物的這種獨特作用,形象地比喻成“二氧化碳泵”(如圖)。同C3植物相比,C4植物大大提高了固定CO2的能力。在乾旱的條件下,綠色植物的氣孔關閉。這時,C4植物能夠利用葉片內細胞間隙中含量很低的CO2進行光合作用,而C3植物則不能。這就是C4植物比C3植物具有較強光合作用的原因之一。
