共找到4條詞條名為受精的結果 展開
- 卵子和精子融合的過程
- 受孕
- 高等維管植物的特有現象
- 2011年美國電影
受精
卵子和精子融合的過程
受精是卵子和精子融合為一個合子的過程。
它是有性生殖的基本特徵,普遍存在於動植物界,但人們通常提到最多的是指的動物。動物受精在細胞水平上,受精過程包括卵子激活、調整和兩性原核融合3個主要階段。激活可視為個體發育的起點,主要表現為卵質膜通透性的改變,皮質顆粒外排,受精膜形成等;調整發生在激活之後,是確保受精卵正常分裂所必需的卵內的先行變化;兩性原核融合起保證雙親遺傳的作用,並恢復雙倍體,受精不僅啟動DNA的複製,而且激活卵內的mRNA、rRNA等遺傳信息,合成出胚胎髮育所需要的蛋白質。
受精后6~7日晚期胚泡透明帶消失后逐漸埋入並被子宮內膜覆蓋的過程,稱受精卵著床。需要經歷三個過程:1.定位;2.黏附;3.穿透。著床后迅速發生蛻變。
動物的精子不像低等植物如苔蘚植物的精子有明顯的趨化性,而是靠自身主動運動或依靠生殖道上皮細胞的纖毛運動抵達卵子附近。人類受精時間:排卵后24小時內。受精地點:輸卵管壺腹部。
已知許多哺乳動物精子經過雌性生殖道或穿越卵丘時,包裹精子的外源蛋白質被清除,精子質膜的理化和生物學特性發生變化,使精子獲能而參與受精過程。
哺乳動物的獲能精子接觸卵周的卵膜或透明帶時,特異地與卵膜上的某種糖蛋白結合,激發精子產生頂體反應:頂體外圍的部分質膜消失,頂體外膜內陷、囊泡化,頂體內含物包括一些水解酶外逸。頂體反應有助於精子進一步穿越卵膜。在海膽卵上,激起精子頂體反應的是卵周膠膜中的某種多糖物質。絕大部分卵的外周都有卵膜,各種卵膜厚度不一,主要組分是粘蛋白或粘多糖;只有少數是裸卵,如腔腸動物的卵。
精子穿越卵膜時,出現先粘著后結合的過程。前者為疏鬆附著,不受外界溫度干擾,無種的專一性,粘著期間,頂體內膜上的原頂體蛋白轉化為頂體蛋白,頂體蛋白有加速精子穿越卵膜的作用;後者是牢固的結合,能被低溫干擾,具有種的專一性。在海膽精子質膜上已分離到一種能與卵膜糖蛋白專一結合的蛋白質,稱作結合蛋白,分子量約30000。
在排卵時處於第二次成熟分裂中期的次級卵母細胞連同周圍的透明帶和放射冠,由於輸卵管上皮細胞纖毛的擺動和肌層的收縮,迅速通過腹腔進入輸卵管壺腹部。如果卵細胞未能與精子相遇,一般在12—24小時內開始變性死亡。
精子一旦與卵子接觸,卵子本身也發生一系列的激活變化。在哺乳動物卵上,則表現為皮層反應,卵質膜反應和透明帶反應,從而起到阻斷多精受精和激發卵進一步發育的作用。皮層反應發生在精卵細胞融合之際,自融合點開始,皮質顆粒破裂,其內含物外排,由此波及整個卵子的皮層。卵質膜反應是卵質與皮質顆粒包膜的重組過程。透明帶反應為皮質顆粒外排物與透明帶一起形成受精膜的過程,卵膜與質膜分離,透明帶中精子受體消失,透明帶硬化。
只有發生頂體反應的精子才能與卵融合。在頂體酶的作用下,精子穿過放射冠,並與透明帶上精子受體糖蛋白分子ZP3相作用,使精子釋放頂體酶,穿過透明帶進入卵周隙。受精開始時,人精子頭側面赤道部的包膜與卵細胞膜接觸,隨即精子的細胞核和細胞質進入卵內。精子進入卵子后,卵子淺層細胞質內的皮質顆粒立即釋放其內容物到膜周圍間隙中,引起了透明帶中ZP3糖蛋白分子變化,使透明帶失去接受精子穿越的功能。與此同時,隨著皮質顆粒的膜與卵細胞融合,使細胞表面負電荷隨之增多,從而制止精子質膜與卵膜的融合,稱之為皮質反應(cortical reaction)。透明帶結構發生變化,稱為透明帶反應(zona reaction)。此時,透明帶對精子的結合能力降低,防止了多精受精(polyspermy)的發生,保證了人類單精受精(monospermy)的生物學特性。
皮質顆粒的內容物(一種胰酶樣蛋白酶)除能破壞或滅活透明帶上與精子結合的受體,還能使透明帶中肽鏈間的交聯增加,對頂體蛋白酶的敏感性減弱,制止精子穿透。通常情況下,雖然有數個精子穿越透明帶,但只有一個精子進入卵細胞內,使之受精。在異常情況下,可以有兩個精子參與受精,即雙精受精。兩個精子同時進入卵子形成三倍體細胞的胚胎,此種胚胎均流產或出生后很快死亡。精子入卵后,卵子迅速完成第二次成熟分裂,此時的精子和卵的細胞核分別稱為雄原核和雌原核。兩個原核逐漸靠攏,核膜消失,染色體融合,形成二倍體的受精卵。
精卵細胞融合時首先可以看到卵子表面的微絨毛包圍精子,可能起定向作用;隨即卵質膜與精子頂體後區的質膜融合。許多動物的精子頭部進入卵子細胞質后即旋轉180°,精子的中段與頭部一起轉動,以致中心粒朝向卵中央。接著雄性原核逐漸形成,與此同時中心粒四周產生星光,雄性原核連同星光一起遷向雌性原核。精子中段和尾部不久退化和被吸收。卵子細胞核在完成兩次成熟分裂之後,形成雌性原核。雌、雄兩原核相遇,或融合,即兩核膜融合成一個;或聯合,兩核並列,核膜消失,僅染色體組合在一起,以建立合子染色體組,受精至此完成。
卵受精之前,代謝水平很低,無DNA的合成活動,RNA和蛋白質的合成都極少。因此排出的卵子,如果未受精,很快就夭折。
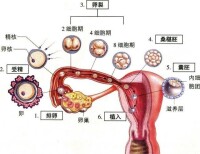
體內由排卵、受精、到著床的過程
在海膽卵激活的早期階段,質膜對鈉離子的通透性增加,鈉離子大量內流,致使質膜在數秒鐘之內去極化;鈣離子自細胞庫存中釋放,使20~30秒鐘之內卵內遊離鈣離子量迅速增加多達100倍;隨著鈉離子內流,氫離子外流,致使一分鐘之內卵pH值明顯增加。這些離子的變化,誘發皮層反應,導致阻斷多餘精子入卵,並激起卵的進一步發有。卵內遊離鈣離子的增加,趕到激活卵內鈣調素的作用,由此進一步激活卵內其他的蛋白質。隨即出現蛋白質合成量的增加,DNA也開始複製。
在海膽卵上,鈣離子也可能是通過鈣調素,激活卵質膜上的某些專一轉運蛋白質,使氫離子向細胞外輸出。卵內pH值的增加,會引起蛋白質合成速率增加和DNA的複製。這種氫離子的外流依賴於鈉離子的內流。這些蛋白質的合成並不依賴於RNA的新合成,而是預存卵中mRNA的去掩蓋以及核糖體激活的結果。
受精機制的研究,是人類有效掌握和控制有性生殖動物繁殖和育種的基本保證之一。人類“試管嬰兒”的誕生只是少數成功的事例。由於受精和進一步正常發育機制方面尚存在許多懸而未決的問題,許多動物的體外受精的嘗試均遭失敗。這些失敗表明基礎理論研究的重要。
凡在雌、雄親體交配時,精子從雄體傳遞到雌體的生殖道,逐漸抵達受精地點(如子宮或輸卵管),在那裡精卵相遇而融合的,稱體內受精。
多發生在高等動物如爬行類、鳥類、哺乳類、某些軟體動物、昆蟲以及某些魚類和少數兩棲類。
凡精子和卵子同時排出體外,在雌體產孔附近或在水中受精的,稱體外受精。水生動物的普遍生殖方式,如某些魚類和部分兩棲類等。
體外受精
通如果因為卵子的成熟程度不適當等原因,而有一個以上的精子進入這類卵子,即所謂的病理性多精受精,則卵裂不正常,胚胎畸形發育,遲早必歸天殤。有些卵子在正常受精情況下,可以有一個以上的精子進入卵子,但只有一個精子的雄性原核能與卵子的雌性原核結合,成為合子的細胞核,其餘的精子逐漸退化消失,稱生理性多精受精。如昆蟲、軟體動物、軟骨魚、有尾兩棲類、爬行類和鳥類的受精。
1875年
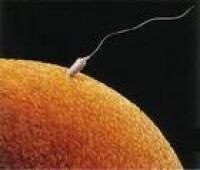
精子到達卵子
20世紀以來
受精研究轉向探討兩性配子結合的機制。美國學者F.R·利利根據沙蠶和海膽上的研究,首先指出卵子分泌出與接受精子有關的物質,他稱之為受精素。40年代前後,另一美國學者A.泰勒就受精素的生物學、化學和免疫學特徵展開了一系列工作,進一步強調卵子成熟過程中排出物對受精的重要意義。與此同時,德國學者M.哈特曼認為在海膽受精過程中,不但卵子能排出雌配素,精子也能排出雄配素,兩者相互抗衡的程度決定著受精成功與否。不久,在兩棲類上,發現卵外膠膜在受精中的作用。
1956年
中國實驗細胞學家朱洗等根據中華大蟾蜍的實驗,提出輸卵管分泌的卵外膠膜,為雌雄配子實現受精所必需。在哺乳動物方面。
1951
張明覺和C.R.奧斯汀分別同時提出精子必須在雌體生殖道逗留一段時間,獲得穿入卵子的能力——獲能,才能有效地使卵子受精。精子獲能的發現使人們找到過去哺乳類卵子離體受精不成功的原因,從而把高等哺乳動物和人類卵子受精的研究推向一個新階段。
精子和卵子為何個體相差懸殊
數量不同
據《中國大百科全書》醫學卷所載,人的卵子肉眼可見,直徑約0.2毫米,是人體最大的細胞,而精子即便以長度計也只有0.06毫米,若以體積比較則只有卵子的千分之一。由於這層關係它們露面的機會也相去甚遠,卵子大約每一個月才排出一個,而不受時間限制的精子一次可排出2億之眾。
卵細胞同樣肩負生殖使命,它們的個體為何相差如此懸殊呢?首先要弄清為什麼動物只分為兩種性別。或許有人會說,是為了限制擇偶時的成功率,維持穩定的數量。可又為什麼讓精子處在被選擇位置呢?這是進化生物學領域的一個大課題。對性有多種定義方法,這裡講的是有性繁殖的生物,具有產生卵子、子房等這類大配子功能的個體稱作雌性,而那些只產生精子、花粉等小配子的則稱其為雄性。
游弋的精子產生什麼樣的配子繁殖成功率高,而且能提高結合之後的子代成活率?通過計算可以找出答案,這種方式出現於20世紀70年代,人們稱其為“遊戲法則”。通俗地解釋就是:養分付出后若全部生成小配子確實可大量產生子代,但是小配子之間結合,造成個體養分缺乏,存活困難。反過來,若全部生成大個體配子,生存率雖然可以提高,而子代數量也明顯減少了。為了解決這一矛盾,通常情況下由一種性別充當配子,用來大量產生可滿足基本功能的最小個體;另一性別以較大個體少量生成,朝這一方向進化下去的結果就造成了大卵子,小精子。
精卵相遇
當卵細胞和精子相遇時另外,小個體有較好運動能力,可以盡情地去尋找能容它們寄生的大個體,這樣的配子有利於生存。於是兩個不同個體形成雌雄兩性,並以集團出現。到了上世紀90年代,英國牛津大學的哈米爾頓教授提出的細胞質非對稱性,又對此做出了新的解釋,他認為這種個體差異是為了避免卵子和精子間鬧“糾紛”,具體原因主要在精子身上,因為它們失去了以線粒體DNA為首的細胞質遺傳基因。如果卵子精子都有細胞質遺傳基因,結合以後就會互相猛攻對方的遺傳基因,為避免這種混亂,結合之前,精子就很大度地事先解除了自己的細胞質遺傳基因,而只帶有核遺傳基因。
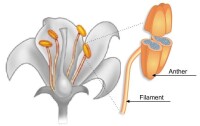
被子植物雙受精
有三種:①同配生殖(同型配子配合);②異配生殖(異型配子配合);③卵式生殖(精卵配合)。
受精的三種形式反映了融合的兩種配子從很少分化到有明顯的分化的進化過程。
融合的兩種配子在形態和大小上是完全相似的,其中有兩種情況:①同宗的個體(由一個個體經無性生殖或營養生殖所產生的後代)所產生的配子可以互相配合,叫做同宗配合;②甲宗的配子只能與乙宗的配合,叫做異宗配合。後者配子雖然大小、形態相同,但在生理上有差別。異宗配合的兩種配子和產生它們的植物體通常分別用正號“+”和負號“-”表示。綠藻中衣藻屬的某些種是異宗配合的。這些種的配子有“+”和“-”不同的交配型,只有不同的交配型的配子之間才能發生凝集和進一步的融合。真菌中的黑根霉以及毛霉屬的一些種也可見到異宗配合生殖。當“+”系和“-”系菌絲相遇,在相對的位置可以產生原配子囊,而後相對的不同交配型的配子囊的原生質體融合。
融合的兩種配子在大小上有固定的差別,這是性細胞進一步分化的表現。例如綠藻中的海松屬,在一個重複分叉的個體上產生兩種配子囊,在一種配子囊中產生具有二根鞭毛的形體較大的配子,另一種配子囊中產生與前者形態相似而形體小得多的配子,大的稱為雌配子,小的稱為雄配子。
融合的兩種配子在結構、能動性和大小上都有顯著的差別。雄配子通常稱之為精子,植物界許多類群的精子是具鞭毛和有運動能力的。精子的核大而細胞質少;高度分化的雌配子稱為卵或卵球,卵細胞的細胞質極為豐富。從低等的藻類、真菌到種子植物都有卵式生殖,而同配生殖、異配生殖只發生在藻類和菌類。在同型配子配合和異型配子配合的受精方式中,性細胞都具有鞭毛,融合的兩種配子可在水中遊動相遇並實現受精。在卵式生殖中,受精的雌配子一般是不動細胞,性器官的結構也較複雜。受精的完成有多種不同的方式。團藻屬的精卵配合是卵式生殖中最簡單的代表。
在團藻的多細胞個體中可同時形成雄性和雌性兩種配子。由一個雄性原始細胞經多次縱分裂產生許多細長而具兩根鞭毛的精子;卵則是由一個雌性原始細胞增大而成,成熟時為球形,無鞭毛,遊動的精子游至被膠質鞘包被的卵中,由一個精子與卵融合完成受精作用。
藻類和真菌植物
藻類和真菌植物還有其他精卵融合的方式。例如真菌中的水霉屬,雌性器官——卵囊在菌絲的末端形成,在卵囊中發育出一至多個卵;產生精子的精子器在卵囊的附近產生,精子數量多,不能遊動。在受精時,由精子器長出的受精管穿入卵囊並將精子射出達到卵旁而實現受精。綠藻中無隔藻屬的性器官與水霉基本相似,但精子能動,成熟的卵囊中只有一個卵。精子從精子器釋放出來以後,通過卵囊喙狀開口進入卵囊與卵融合。
在苔蘚和蕨類植物中,精子和卵都包藏在多細胞的性器官中,受精還不能離開水的條件。地錢的雌性器官形如瓶狀,稱為頸卵器。頸卵器與精子器分別生於葉狀配子體的托上。地錢是雌雄異體的。在頸卵器中只產生一個卵,而在精子器中產生許多長形捲曲而具二根鞭毛的精子。當精子器成熟時,精子釋放出來,如果植物體背面有水,精子可以游至頸卵器與卵實現受精。蕨類植物的頸卵器和精子器是在同一個配子體的下表面產生,能遊動的精子經配子體表面的水層游進頸卵器內。
種子植物生殖器官的結構更為複雜,在有性生殖過程中產生了花粉管,藉助花粉管,受精不再依賴水的條件,這是種子植物適應陸地生活的一個重要因素。產生雌配子的雌配子體在大孢子囊即胚珠的珠心處發育。裸子植物中的大多數仍保留頸卵器的結構,卵在頸卵器內形成。在小孢子囊中發育的花粉粒,即前期的雄配子體。在雄配子體中形成兩個精子。精子多不具鞭毛,但在蘇鐵目和銀杏目仍具遊動的精子。被子植物的雌配子體稱為胚囊,卵直接在其中產生;精子完全失去鞭毛。這些特點決定了種子植物的受精必先經過傳粉。裸子植物的胚珠是裸露的,經過傳粉,花粉被送到胚珠的珠孔處,而在被子植物中,胚珠著生在雌蕊的子房內,花粉粒不能直達胚珠而是落在柱頭上,再由萌發生長的花粉管將精子輸送到雌配子附近。在裸子植物中,進入頸卵器的兩個精子只有一個與卵融合。但在被子植物中進入胚囊的兩個精子則分別與卵和極核融合。這就是被子植物特有的雙受精。
受精過程中識別作用
20世紀40年代已認識到衣藻的有性生殖是由一種稱為配素的物質所控制。後來人工培養的試驗進一步明確配子狀態的細胞釋放至培養基的物質能引起相對的交配型的配子凝集;即從“+”系的細胞釋放的物質可使“-”系的細胞凝集,反之,從“-”細胞釋放的物質可凝集“+”細胞,在同性凝集之後,並不發生細胞融合。這種物質稱為同種凝集素,它是一種含糖蛋白的物質,存在於鞭毛的表面的微小泡中,它不僅有性的專一性,而且有種的專一性,無疑是涉及配子的粘著和識別反應的物質。
在真菌的毛霉目的有性生殖中存在類似的識別反應。例如毛霉屬異宗配合時是通過“+”系和“-”系配子囊的融合,人工培養的實驗已經證明,控制異宗配子囊的融合分兩步。首先要由三孢酸誘導產生兩種交配型的原配子囊,而三孢酸只有在兩種交配型存在的情況下才能形成,因為一種交配型產生的前體只能被相對交配型轉變為三孢酸,即“+”系產生的前體只能被“-”系轉變為三孢酸,“-”系產生的另一前體只能被“+”系轉變為三孢酸。然後不同交配型的原配子囊相遇才發生融合。這一識別反應,推測是由於在配子囊頂端區域的壁上存在一種含糖蛋白的識別物質。
苔蘚和蕨類植物的卵包藏在頸卵器中,遊動的精子到達頸卵器和卵結合必需游至一定距離,精子能“發現”卵,顯然是存在一定的相互作用的物質。在這兩大類群的植物中,精子的向化性已被肯定。向化性指精子趨向卵或頸卵器產生的化學物質。W.F.P.普菲費爾最早研究歐洲蕨(Pteridium aquilinum)的精子向化性的實驗證明,由頸卵器產生的L-蘋果酸是有效的吸引劑。它將含有自來水的(包含10 %瓊脂) 10%L-蘋果酸鈉的滴管插入精子的懸液中,蘋果酸鹽離子立刻從滴管中有級差地擴散到四周的介質中,精子在一秒鐘內便移向滴管的口,即向蘋果酸鹽來源的方向移動。在缺少蘋果酸鹽的自來水中,精子是隨意行動的;如果在均質的蘋果酸鹽的溶液中,即在級差不存在時,精子在溶液中也保持均勻的分佈。在問荊曾試驗過12種有機酸,精子僅對L-蘋果酸和酒石酸敏感。
被子植物
在被子植物中,精子由花粉管傳送至胚囊。對花粉管通過花柱定向生長的原因,曾有各種解釋。有的推測也是向化性因素的控制,即認為鈣是吸引花粉管生長的因素;有的認為花粉管早期生長受營養核中DNA轉錄的mRNA所控制;還有的假設花粉管定向生長決定於柱頭至子房電位差等等。這是一個尚無定論的問題。
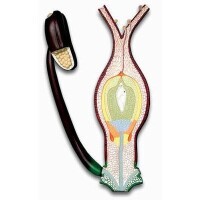
被子植物受精模型
落到柱頭上的花粉是被“接受”或是被“拒絕”決定於它們之間的相互識別。如果是親和的,植物的花粉粒便在柱頭上萌發出花粉管長入柱頭,並沿一定的途徑通過花柱到胚囊,在那裡釋放出精子;如果植物的花粉與柱頭是不親和的,雌蕊將抑制花粉的萌發或花粉管的生長而阻礙受精。
植物受精的不親和性是由兩親本的基因型所決定,並表現出生理上的抑制反應。自交不親和性的生理基礎根據對十字花科和菊科的一些植物的研究,認為是花粉與雌蕊組織間識別反應的結果。與識別有關的物質,在花粉的一方是存在於花粉壁的蛋白質;在雌蕊一方已確定的是覆蓋在柱頭表面的蛋白質薄膜。
花粉有外壁和內壁。組成外壁的物質是類胡蘿蔔素和類胡蘿蔔酯氧化多聚化而形成的孢粉素。在外壁物質所形成的結構之間有由絨氈層合成和運入的蛋白質,當內壁發育沉積果膠纖維素時,由花粉本身細胞質合成的蛋白質併入其中。這樣,當花粉成熟時,其外壁有孢子體(絨氈層細胞)基因型控制產生的蛋白質;內壁有配子體(花粉中細胞)基因型控制產生的蛋白質,這些都是起識別作用的蛋白質。
關於雌蕊對花粉的識別方面,如今對干型柱頭(柱頭成熟時無液體分泌物)的認識比較清楚。例如在十字花科和菊科的植物,柱頭的表面產生乳頭狀單細胞,它具果膠纖維素的壁及一層不連續的角質層。乳突狀細胞在發育過程中,通過壁和角質層還分泌一些物質形成一層薄膜覆蓋在表面,此薄膜主要成分是蛋白質,它不僅提供了一個有利於“捕捉”花粉的粘的表面,而且具有識別“感受器”的特點。當與花粉壁蛋白結合後接著發生“接受”或“拒絕”反應。
花粉落在柱頭上,從柱頭吸水膨大,在接觸后的幾分鐘內,花粉壁蛋白質即釋放出來與柱頭表面的蛋白質薄膜結合,如果是親和的結合,花粉萌發,由於酶的作用,柱頭的角質層被溶解,花粉管進入柱頭並在花柱中順利地生長。在不親和的情況下花粉管生長停滯,或不能進入柱頭(孢子體系統控制),或花粉管在花柱中生長停頓和破裂(配子體系統控制)。蘿蔔、白菜、大波斯菊和向日葵等植物在不親和的自交中,傳粉幾小時后,柱頭的乳突狀細胞中即產生胼胝質的凸透鏡狀結構;而在親和的交配中則沒有胼胝質反應。這種胼胝質反應可以作為在一定親緣關係範圍內檢驗親和性的指標。一種花粉落到柱頭上能夠萌發,並同時引起柱頭乳突狀細胞的胼胝質形成(禾本科植物在花粉管頂部形成胼胝質)這是不親和性的反應。種內不親和與種間不親和都發生這種反應,這種反應標誌了兩親本之間尚有一定的親緣關係;如果花粉既不萌發,又不引起乳突細胞呈現胼胝質反應,則反映了兩親本之間無親緣關係的不可雜交性,這常是不同科間的遠緣雜交表現的情況。
種間雜交
有一些實驗證明花粉與柱頭之間存在識別作用。銀白楊和加拿大白楊的種間雜交通常是不育的,異己的花粉停滯在柱頭上。但從銀白楊的花粉提取出其表面的蛋白質與不親和的加拿大白楊的花粉一起加在銀白楊柱頭上,則可獲得一定數量的雜交種子;用殺死的銀白楊花粉與加拿大白楊花粉混合后授與銀白楊,也得到同樣的結果。這表明母本花粉的壁蛋白與柱頭的識別作用,可使不親和的異己花粉被接受。這樣的親和花粉與異己花粉混合授粉的方法在生產上已作為產生雜種的一種手段。
離體傳粉和受精
20世紀60年代已可使被子植物帶胎座的胚珠、整個雌蕊或子房置於離體培養的條件下實現傳粉和受精。
1962年K.甘達及其合作者報道了他們用罌粟為材料在無菌條件下,將沒有受精的帶胎座的罌粟胚珠,置於裝有消毒的營養培養基的試管中,同時用無菌針從花藥中取出花粉撒在胚珠的表面上,結果發生了與正常相似的受精作用,最後結出有生活力的種子。這些種子可在試管中長成健康的幼苗,這是第一個離體傳粉和受精成功的實驗。迄今已報道有40多種植物獲得成功,主要屬罌粟科(如薊罌、虞美人)、茄科(如煙草、矮牽牛)、石竹科(如麝香石竹、異株女婁菜)及十字花科(如甘藍)的植物,近年來又在禾本科的玉米和小麥中獲得成功。
離體傳粉
離體傳粉和受精的技術在育種過程中有可能作為克服有性生殖中自交或雜交存在的不親和性障礙的手段。自交不親和植物在受精過程中的障礙,通常發生在柱頭或花柱中,即由於花粉與柱頭識別反應導致花粉管生長受到抑制,不能進入柱頭或在花柱中生長停滯以致受精失敗。因此有可能利用離體的胚珠授粉順利完成受精。這方面已在矮牽牛屬植物試驗成功。例如自交不親和的腋花矮牽牛用一個子房的胎座上的全部胚珠進行離體培養和授以自花花粉。結果花粉管生長正常,最後形成具正常發育的胚和胚乳的種子。由離體受精得到的種子長成的第一代植株象它的親本一樣,是二倍體的,能育和自交不親和的。另一方面,曾在石竹科、罌粟科、茄科及禾本科的一些植物試用離體傳粉和受精的技術克服雜交不親和性。其中許多雜交組合雖然能達到受精,但未能獲得有生活力的種子。不過也有少數得到雜種種子和幼苗的。如:①栽培煙草(2n=28)與德氏煙草或與蓮座煙草(2n=40)雜交,這是常規的雜交方法難以成功的,然而,用離體傳粉的方法,在試管里得到了雜種種子而且種子在培養基上萌發成幼苗;②玉米與墨西哥玉米在試管中雜交,獲得了具發育良好的胚和胚乳的種子;③小麥(2n=42)的子房在離體培養條件下授以黑麥(2n=14)的花粉,得到具有器官分化的胚和含胚乳的穎果。將這種雜種胚從穎果中解剖出來進行胚培養,結果長成雜種幼苗。幼苗根尖染色體數為小麥和黑麥體細胞染色體半數之和,即28(21+7)個。
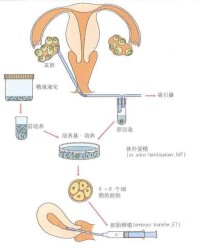
人工受精的條件
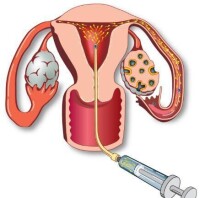
人工受精過程圖解

基本信息
- 中文名
- 受精
- 外文名
- fertilization
- 概念
- 卵子和精子融合為一個合子的過程
- 起始時間
- 1875年
- 意義
- 產生新的生命
- 代表科學家
- 赫特維希