第三紀
第三紀
第三紀(TertiaryPeriod),新生代的最老的一個紀(距今6500萬年~距今180萬年),第三紀的重要生物類別是被子植物、哺乳動物、鳥類、真骨魚類、雙殼類、腹足類、有孔蟲等,這與中生代的生物界面貌迥異,標誌著“現代生物時代”的來臨。
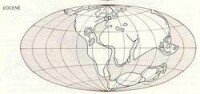
第三紀時期全球的陸地分佈
1833年,英國C.萊伊爾在研究法國巴黎盆地軟體動物化石時,發現地層越新,軟體動物與現代種屬相同的越多。他根據地層中含有現代種屬的百分比,將第三紀劃分為始新世、中新世和上新世。
1854年,E.貝利希在德國發現早於中新世的沉積物,提出漸新世。1853年,M.赫奈斯依生物群和沉積物的相似性,將中新世和上新世合稱晚第三紀。
1866年,K.F.瑙曼則把漸新世和始新世合稱早第三紀。1874年,W.P.夏姆珀根據巴黎盆地發現的植物化石,他認為這些化石的層位早於始新世,而稱之為古新世。因此,第三紀劃分為早第三紀和晚第三紀。早第三紀包括古新世、始新世和漸新世,晚第三紀包括中新世和上新世。早、晚第三紀的分界線約在2330萬年前。

第三紀遺留的地貌
的名稱是根據現代海洋無脊椎生物種屬在第三紀化石中占的比例命名的。因此,根據提出這個系統的查爾斯·萊爾爵士的意見,始新世(意思是“現代的拂曉”)有比更新世(譯為更為現代)較少的現代物種。在另一種分類中,將第三紀看作一個代,其中的前三個世構成老第三紀而後兩個構成新第三紀。
脊椎動物的變化主要表現在爬行動物的衰亡,哺乳類、鳥類和真骨魚類取而代之,興起且高度繁盛。第三紀的早期,仍生活著古老、原始的哺乳動物;到了中期,現代哺乳動物的祖先先後出現,逐漸代替了古老、原始的哺乳動物;第三紀晚期,現代哺乳動物群逐漸形成,更是偶蹄類和長鼻類繁盛的時期。尤其馬的進化很快。
中生代末,海生無脊椎動物有明顯的興衰現象。盛極一時的菊石類完全絕滅,箭石類極度衰退,而雙殼類、腹足類、有孔蟲、六射珊瑚、海膽、苔蘚蟲等則進一步繁盛。第三紀出現的有孔蟲分佈廣泛、進化迅速,對於海相第三系的劃分與對比很有意義。此外,它們的生態分帶已應用於確定沉積盆地水深的變化。原生動物中的放射蟲在第三紀也十分繁盛,在深海研究中佔有突出地位。雙殼類在第三紀有很大發展,腹足類在第三紀進入極盛期。
陸生的無脊椎動物以雙殼類、腹足類、介形類為主,可以根據它們不同時期組合面貌的變化,進行陸相第三系的劃分。
(開始於6500萬年前,結束於2330萬年前)

第三紀化石
豐富的各種生物類群的。這些化石為下第三系的劃分、對比以及標準層序和層型剖面的建立起著關鍵性的作用。世界上不少地區在早第三紀形成了界線較為分明的海相和海陸過渡相的地層,如西歐下第三系標準層序劃分為11個階。在北美中西部、亞洲中部等地區也形成了以河湖相為主的陸相沉積,尤以美國落基山脈的盆地分佈區最為典型。
下第三系除以岩石地層、古地磁和同位素測定為依據外,主要採用古生物組合(或動物群)、層序和層型剖面等對比方法來劃分。由於各門類生物進化速率不同,各地沉積並非同時,因此地層的劃分常與上述研究提供的論證不相吻合,以致對比和界線常引起爭議。如海相地層中的達寧階(丹麥)和阿基坦階(法國)的時代歸屬就有不同認識。
有人認為達寧階的棘皮動物(Echioids)等具有晚白堊紀的特徵,但是據克里米亞、高加索和裏海周圍地區以及丹麥、瑞典南部浮遊有孔蟲和棘皮動物化石的研究,它們與古新世類型有密切關係。因此較多的學者認為,中生界與新生界分界應置於馬斯特里赫階與達寧階之間。但仍有一些學者認為,中生界與新生界界線應劃在達寧階與蒙蒂階之間。阿基坦階的歸屬問題,古生物學家從軟體動物和哺乳動物研究認為,它的生物群與下部斯坦普階的化石有著明顯的繼承關係,而與晚第三紀相比有一定的差異,他們主張阿基坦階定為漸新世的頂界。
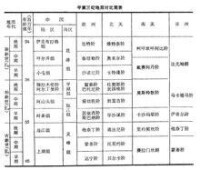
早第三紀地層對比簡表
不多,主要以美國居多。亞洲在20世紀初僅在蒙古發現了一個地點。1960年以來,中國在8個省和自治區陸續發現了不少重要的古新統地點。中國陸相古新統以下部的上湖組和上部的池江組為代表。古新統已發現的動物化石几百種,其中含有脊椎動物100多種。始新統在中國有良好的發育,如內蒙古地區的典型層序阿爾丁曼階和錫拉木侖階已獲國際上公認。
中國海相下第三系見於喜馬拉雅山區、塔里木西緣和東海陸架──台灣島。喜馬拉雅山區有特提斯型古新統和始新統,古新統包括宗浦組下部,下、中始新統包括宗浦組上部和遮普惹組,它們含大有孔蟲、介形蟲和軟體動物化石。塔里木西緣下第三系主要是喀什群,包括古新統、始新統,另外,漸新統包含於烏恰群底部,它們含有孔蟲和軟體動物等化石,屬於中亞副特提斯海區。台灣北港地區鑽孔中發現古新統雲林組,有鈣質超微化石,西部山麓帶和中央山脈帶有始新世畢祿山組、十八重溪組、達見組,晚始新世―早漸新世西村組,早漸新世四棱組、中、晚漸新世水長流組、花港組和五指山組,它們主要以有孔蟲、鈣質超微化石和軟體動物化石為時代依據。東海陸架下第三系發育良好,其中古新世靈峰組、始新世甌江組有孔蟲、鈣質超微化石序列完整,為西太平洋地區最完善的海相地層序列。
此外,為了使典型層序逐步完善和具有一定的代表性,仍需加強界線和全球性事件的研究。早第三紀地層對比簡表(見表[早第三紀地層對比簡表]),其中有些地層單元的對比關係尚不夠了解,如海相與陸相有關層位的對比等,這在闡明區域或國際間有關層位相互關係帶來一定的困難。
(距今2330萬年—距今164萬年)晚第三紀地層的劃分與對比有其獨特而複雜的歷史。
統的劃分

第三紀化石
地中海周圍海相雙殼類化石組合中現生種的含量確定的:中新世含現生種18%;上新世含現生種49%。萊伊爾定名時,沒有指定典型層位和地點,也沒有確定界線,更沒有考慮到它們是否能在全球適用。這是造成後來長期爭論和混亂的根本原因。雖然如此,絕大多數地質學家都同意,上第三系的統的劃分應以研究歷史最久的地中海周圍的海相地層為準。
中新統的底界,即上第三系的底界應以阿基坦階的底界為準,它代表了漸新世末海退之後的一次廣泛海侵的開始,大約發生於2400萬年前。中新統的上限,即上新統的下限應在墨西拿階和贊克爾階之間。墨西拿階代表古地中海區域一次廣泛的海退,是有名的成鹽期。贊克爾階代表一次海侵。這一條界線,由於長期來被錯誤地與陸相地層中的三趾馬動物群的出現聯繫在一起,而推定為距今約1200萬年(這也是古哺乳動物學家根據生物演化規律而推算出的結果)。
如今已有大量的證據證明對海相中、上新統的界線年齡的推斷是錯誤的。實際它距今只有大約550萬年。上新統的頂界,即上第三系的頂界在皮亞琴察階和卡拉布里亞階之間。第18屆國際地質大會(1948)把這條界線定為上第三系與第四系在海相層中的界線。關於這一界線的年代的意見已趨於一致,即生物群總體面貌發生較大變化的界面與古地磁年表中松山反向期中的奧杜威正向亞期的上界大體同時,即距今約170萬年。中、上新統在時限上太不均衡,上新統的時限太短(約300萬年),而中新統的時限又太長(約1900萬年)。此外,這些統的界線不能反映全球性重大的地質和生物事件。事實上,各大陸的晚第三紀的生物和地層年代劃分的界線大多與上述界線不相吻合。因此,這種統(世)的劃分已失去其作為全球性劃分單元的性質。
階的劃分
中國晚第三紀哺乳類層序及與西歐對比

第三紀動物
東歐,上第三系主要以副地中海的海相、半鹹水封閉內陸海和陸相互層為代表。這些地層在19世紀中期和後期曾是著重研究的對象,出現過大量的地層名稱。其中有些是相當著名並被廣泛應用的,例如蓬蒂階等。副地中海有極為複雜的演變歷史。因此,它的地層單位之間以及它們和古地中海海相階的對比造成過極大的混亂。
從20世紀60年代末開始,經過多學科長年大量的工作,這些問題已逐步得到澄清。在地中海晚第三紀地層分委員會第7次(1979,雅典)和第8次(1985,布達佩斯)大會上已經提出了中副地中海和東副地中海兩個區域地層劃分方案,地層對比問題也已基本解決。

遺留地貌
出現的分子等。這兩種劃分已經成為當前陸相地層的全球性對比的標準。海相沉積僅分佈在台灣、海南島及沿海極個別的地段。中國上第三系研究起步晚,從20世紀20年代才開始,而且主要限於晚第三紀中後期的哺乳動物化石方面。在30~40年代,曾發表大量有關三趾馬動物群方面的專著。但在地層劃分方面工作不多。80年代以前,中國一直沒有一個獨立的上第三系階一級的劃分體系;三趾馬動物群和泥河灣動物群分別借用了歐洲的蓬蒂期和維拉弗朗期的名稱;其他動物群只是籠統地劃歸中新世或上新世;而且一直沒有發現早中新世的哺乳動物。
1984年,李傳夔等首次提出一個與歐洲大體對應的7個期(階)的劃分方案。最近的工作表明,中國至少也能劃分出15個哺乳動物帶來。中國下中新統僅發現於青海西寧、甘肅蘭州、河西走廊、新疆準噶爾盆地等處,主要為河湖相碎屑沉積;中中新統發現於寧夏同心、陝西藍田、江蘇泗洪、內蒙古通古爾、山東山旺和雲南開遠等地。大部分屬河湖相沉積,也有和等湖沼沉積。晚中新統廣布全國,只華南和東北尚未發現哺乳動物化石。黃河以北、汾河以西主要為土狀堆積,即所謂的三趾馬紅土,汾河以東為河湖相沉積;黃河與長江之間發現較少,在鄭州和襄樊一帶有灰岩;雲南則多沼澤相,有褐煤。70年代在西藏希夏邦馬峰北坡也發現三趾馬化石。上新統發現較少,僅零散分佈於陝西、山西、河北等省和盆地中。
早第三紀由於海底擴張、古陸解體,對世界上整個地質構造格局、古地理環境產生了重大的變化。早第三紀時,古地中海最終消失,亞洲大陸最後形成,青藏高原的升起,阿爾卑斯山、喜馬拉雅山、落基山和安第斯山等現代山系相繼形成,吐爾蓋海峽和巴拿馬地峽的出現或消失,氣候分帶漸趨明顯,地球上的自然環境向著多樣性發展。

遺留地貌
北非阿特拉斯山區、義大利大部、法國和西班牙的地中海沿岸區都曾為古地中海區,法國的西海岸、北歐地區則被大西洋所佔;在北美西海岸南部以及墨西哥灣濱海區也被海洋所佔。但是,中國的情況不同,中國的東海岸在晚第三紀的大部分時間裡更向東擴展,特別是渤海和黃海,大部分還都是陸地。當然最大的不同是副地中海的存在。副地中海是在阿爾卑斯山脈從水下升起后殘留在山脈以北部分的水體。水體最大時東西長約5000公里,面積為現地中海的一倍半,從法國羅訥河谷開始,沿阿爾卑斯北麓向東延伸,至少延伸至鹹海一帶。除羅訥河谷外,副地中海在不同的時期曾通過南斯拉夫薩格勒布低地與亞得里亞海,通過希臘與愛琴海和通過伊朗西北部與波斯灣及印度洋相連通。在中中新世末,約在1300萬年前,基本與其他海洋隔絕,變成內陸海,幾經分隔,剩下今日的鹹海、裏海、黑海和匈牙利的巴拉頓湖。
晚第三紀是重要的山脈形成期。現今地球上較高的山脈都是這一時期形成的,如歐洲的阿爾卑斯山、非洲的阿特拉斯山,亞洲的喜馬拉雅山,北美的落基山和南美的安第斯山等;一些老的夷平山地,如天山、秦嶺等也有活化抬升現象。地殼運動也引起了普遍的火山噴溢。太平洋東、西兩岸有大量玄武岩流。在中國境內的有漢諾壩玄武岩,東北地區大片出露,山東山旺和浙江嵊縣的玄武岩等。地殼運動也使歐亞大陸與非洲和北美數次分合,形成多次動物群交流。南、北美洲也在上新世末相連過。
晚第三紀開始時,氣候由涼爽向濕熱轉變,中中新世以後,西藏高原的升起無疑對全球的氣候產生重大影響。在中國的表現則是氣候分帶日益明顯,並逐漸轉向乾旱涼爽。

遺留地貌
北半球年平均氣溫曾下降3℃左右;到古新世後期,氣溫一度回升;而始新世晚期氣溫又有下降趨勢,直至以後才逐漸回升到原有水平。在早第三紀晚期,中國氣候有明顯的南北分帶,如南北為兩個潮濕帶,中部為乾旱帶。氣候帶的展布與今日的緯度斜交。晚第三紀開始時,氣候由涼爽向濕熱轉變。中中新世后,西藏高原的抬升對全球氣候產生重大影響。中國的氣候分帶日益明顯,並逐漸轉向乾旱涼爽。
總之,第三紀的氣候比起如今的氣候來,更為溫暖、濕潤而且較少變化。在第三紀早期,熱帶和亞熱帶氣候遠遠延及加拿大北部邊界,在稍後的時期內大平原地區則呈現出乾旱境況。趨近第三紀末,氣候逐漸變冷,預示著更新世最早的冰期即將來臨。
在第三紀,中歐、北非和北美東部墨西哥灣沿岸屬於熱帶氣候,而北非、西亞、中亞和中國華南大部地區,則屬於乾燥氣候帶;東北亞、西西伯利亞、中歐、北美西北部、中國東北地區以及澳大利亞東南部和南美南部,屬於溫涼潮濕氣候帶。

遺留地貌
到早第三紀極度繁盛。以前由古代羊齒和各種松柏組成的植被逐漸為被子植物所替代。這時的被子植物基本上是喬早第三紀是哺乳動物歷史上一個很重要繁衍和進化時期。在晚白堊世時,哺乳動物共有10多個科,但古新世時卻增加到40多個科。這些哺乳動物除白堊紀已有的多瘤齒獸目、食蟲目、目和踝節目外,更重要的是各種古老和土著類型的有胎盤類大量發展和分化。它們絕大部分與現代哺乳動物各目都沒有直接的系統關係;有許多種類是離奇的或特化的。始新世的哺乳動物科一級的總數比古新世約增加80%;奇蹄目和偶蹄目兩大現代目是動物群中的重要角色。
此外,嚙形動物、食肉目也有了較大的發展和繁衍。漸新世哺乳動物群具有更近代化的面貌。它顯示了過渡的特點,一方面是“古老”或“土著”類群的大量衰退、絕滅;另外是近代類群的興起和開始發展。其他脊椎動物如軟骨魚類、硬骨魚類、鱷、龜鱉類和蜥蜴類在早第三紀也是常見的化石;鳥類化石也時有發現。至於無脊椎動物,其中雙殼類如牡蠣、蚶、海扇和珠蚌異常繁盛;腹足類屬種繁多,分佈廣泛;介形類主要有浪花介、土菱子介、金星介和達爾文介等。葉肢介屬種單調,主要是雕飾葉肢介、狹葉肢介等,已處於衰落時期;有孔蟲和棘皮動物在世界上分佈很廣,大有孔蟲主要有圓盤蟲類、貨幣蟲類等。由於古地中海區海相早第三系中廣布大型原生動物貨幣蟲,故在歐洲常稱早第三紀為貨幣蟲紀。而昆蟲化石群基本面貌與近代昆蟲相似。

第三紀動物群示意圖
足類和介形類大量衍生。海洋中各種原生動物,如有孔蟲、放射蟲等極為繁盛,在海相地層劃分中起著非常重要的作用。哺乳動物中,歐亞大陸早第三紀的特徵門類,如有袋類、肉齒類,奇蹄目中的雷獸、原始貘類、兩棲犀、巨犀,偶蹄目中的石炭獸、古豬獸等,除少數殘存至中新世初期外,均相繼滅絕。代之而起的是長鼻目、肉食目中的鬣狗科、熊科、奇蹄目中的安琪馬和三趾馬,犀中的板齒犀、大唇犀,偶蹄目中的豬類,反芻動物中的鹿、羊、長頸鹿,以及嚙齒目、兔形目中的大量屬種。其中絕大部分都是滅絕的種和屬。根據哺乳動物的狀況,晚第三紀可劃分為4個時期:早中新世是殘存的和高度特化的早第三紀分子和少量晚第三紀分子的時期;中中新世,是安琪馬動物群時期,此時長鼻目自非洲、安琪馬自北美遷入歐亞形成一個全新的動物群;晚中新世至早上新世,為三趾馬動物群時期,三趾馬從北美遷入,大批草原型動物大量出現;晚上新世,真象從非洲,真馬從北美遷入歐亞大陸形成另一個動物群,並逐步演化為今日之動物群。
非洲的動物群和歐亞大陸的很接近,只是某些類別的祖先類型(長鼻類、蹄兔類等)出現更早,而有些門類則延續時間更長(如肉齒類等)。北美和歐亞大陸只是間斷聯接,動物群差別較大,屬不同動物區系。至於南美洲和大洋洲,差別就更大了。

第三紀動物
島、中東、蘇聯、加利福尼亞和路易斯安那州與得克薩斯州的墨西哥灣沿岸地區都產出大量的石油。墨西哥灣沿岸平原的鹽丘產出岩鹽,也產出石油。在蒙大拿州、懷俄明州、俄勒岡州及華盛頓州都有在第三系中採煤的礦山。第三紀早期(始新世)的綠河組是油頁岩的重要來源,美國的落基山地區、墨西哥、秘魯、玻利維亞在第三系地層開採銅、金和銀礦床。在馬里蘭州、弗吉尼亞州和加利福尼亞州第三系地層中產出硅藻土。
6徠千5百萬年前):除恐龍外,一些在白堊紀集群滅絕中倖存下來的爬行動物類群仍繼續生活下去,哺乳動物和鳥類保留一定的古老特色,並進一步發展,在哺乳動物中出現早期的馬、大象和熊類。植物界中菊科(Compositae)植物開始出現。
(5千3百萬年前):草本(科)植物(grass vegetation)出現並與豆科(Leguminosae)植物、菊科植物一起繼續繁榮。古老動物群逐漸被現代動物群的祖先替代,在4千萬年前,開始出現草食性動物(grazing)和猴子,部分哺乳動物類群(鯨魚、海豚)重返海洋生活。有孔蟲發展並繁盛。
(3千6百萬年前):最早的猿類出現,大型哺乳動物和鳥類在地球上廣泛分佈,如犀類中出現古今陸上最大的哺乳類動物巨犀。大量陸地被草原復蓋。
(2千3百萬年前):靈長類在中新世佔有重要地位,如森林古猿分佈較廣,到中新世末,類人猿(hominid apes)與大型猿類分開演化,類人猿輻射演化並達到演化的頂點,出現西瓦古猿(Sivapithecusa)——具有現生猿類和人類特徵的類人猿。
(5百萬年前):出現最早的人類--南方古猿(Australopithecus)。
早第三紀由於沉積環境多樣化,在中國不僅形成了巨厚的磨拉石建造,而且在廣大地區形成了各種成因類型的陸相、海相、海陸過渡相的沉積物。這些沉積物中蘊藏的資源相當豐富,主要有含銅砂岩和各種鹽類。生成石油的原始物質是有機物。由於早第三紀有大量動植物遺體的堆積,不僅在海相地層而且在陸相地層中都有油田的存在。石油主要見於長期下沉的拗陷和大中型湖泊沉積盆地,如早第三紀初、中期有關層位中。煤受一定的構造和氣候因素控制,常見於暖溫(亞熱帶)地區,因氣候溫暖潮濕適宜於煤的形成和聚集。早第三紀是一個重要的成煤時期,在中國主要見於秦嶺以北,賀蘭山-六盤山以東地區和南嶺以南珠江-右江地區。同時含煤層位中大都含有油頁岩層,如中國撫順、茂名均見有較厚的油頁岩層。早第三紀鹽類沉積相當豐富,主要有石膏、岩鹽、芒硝、天然鹼和鉀鹽等。鹽類的形成除具備深坳而封閉較好的盆地外,更重要的是要具有較長期乾旱氣候的環境。早第三紀時,中國乾旱帶佔有相當大的地區,是鹽類富集的天然場所。此外,還有含銅砂岩、鈾的氧化物等沉積。
上第三系形成的最主要的礦產是和。地中海和北美東、西海岸的大油田有相當一部分是晚第三紀形成的。地中海地區麥興階是重要的膏鹽層。此外,還有褐煤、殘積型鐵礦、錳礦和硅藻土礦等。

基本信息
- 拼音
- dì sān jì