視覺器官
視覺器官
視覺器官(visual organ)是人和動物利用光的作用感知外界事物的感受器官。光作用於視覺器官,使其感受細胞興奮,其信息經視覺神經系統加工后便產生視覺。通過視覺,人和動物感知外界物體的大小、明暗、顏色、動靜,獲得對機體生存具有重要意義的各種信息,至少有80%以上的外界信息經視覺獲得,視覺是人和動物最重要的感覺。脊椎動物的視覺系統通常包括視網膜,相關的神經通路和神經中樞,以及為實現其功能所必需的各種附屬系統。這些附屬系統主要包括:眼外肌,可使眼球在各方向上運動;眼的屈光系統(角膜、晶體等),保證外界物體在視網膜上形成清晰的圖像。
視覺器官是人和動物利用光的作用感知外界事物的感受器官。光作用於視覺器官,使其感受細胞興奮,其信息經視覺神經系統加工后便產生視覺。通過視覺,人和動物感知外界物體 的大小、明暗、顏色、動靜。
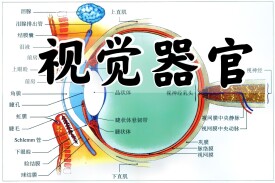
引起視覺的外周感受器官是眼,視覺器官由含有感光細胞的視網膜和作為附屬結構 的折光系統等部分組成。人眼的適宜刺激是波長380-780nm的電磁波;在這個可見光譜的範圍內,人腦通過接受來自視網膜的傳入信息,可以分辨出視網膜像的不同亮度和色澤,因而可以看清視野內發光物體工反光物質的輪廓、形狀、顏色、大小、遠近和表面細節等情況。自然界形形色色的物體以及文字、圖形等形象,通過視覺系統在人腦得到反映。在人腦獲得的全部信息中,大約有95%以上來自視覺系統,因而眼無疑是人體最重要的感覺器官之一。
除了控制眼球運動的眼外肌和起保持、營養作用的鞏膜、脈絡膜等結構外,眼內與視覺傳入信息的產生直接有關的功能結構,是位於眼球正中線上的折光系統和位於眼球後部的視網膜。由角膜經房水、晶狀體、玻璃體直至視網膜的前表面,都是一些透明而無血管分佈的組織,它們構成了眼內的折光系統,使來自眼外的光線發生折射,最後成像在視網膜上,視網膜具有同神經組織類似的複雜結構,其中包含有對光刺激高度敏感的視桿和視錐細胞,能將外界光刺激所包含的視覺信息轉變成為電信號,並在視網膜內進行初步處理,最後以視神經纖維的動作電位的形式傳向大腦。因此,形容眼的功能首先要研究眼內折光系統的不學特性,搞清楚它們怎樣能把不同遠近的物體成像在視網膜上以及形成清晰物像的限度;其次要闡明視網膜是怎樣對視網膜成像進行換能和編碼的。
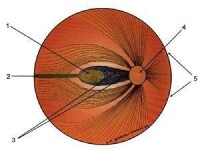
眼和視網膜眼呈球形,由鞏膜所包圍。鞏膜在前方與透明的角膜相接。角膜之後為晶體,相當於照相機的鏡頭,是眼睛的主要屈光系統。在晶體和角膜間的前房和後房包含房水,在晶 體后的整個眼球充滿膠狀的玻璃體,可向眼的各種組織提供營養,也有助於保持眼球的形狀。在眼球的內面緊貼著一層厚度僅0.3毫米的視網膜,這是視覺神經系統的周邊部分。在視網膜與鞏膜之間是布滿血管的脈絡膜,對視網膜起營養作用。
角膜和晶體組成眼的屈光系統,使外界物體在視網膜上形成倒像。角膜的曲率是固定的,但晶體的曲率可經懸韌帶由睫狀肌加以調節。當觀察距離變化時,通過晶體曲率的變化,使整個屈光系統的焦距改變,從而保證外界物體在視網膜上成像清晰。這種功能叫做視覺調節。視覺調節失常時物體即不能在視網膜上清晰成像,可以發生近視或遠視,此時需用合適透鏡來矯正。在角膜與晶體之間,有虹膜形成的瞳孔起著光闌的作用。瞳孔在光照時縮小,在暗處擴大來調節著進入眼的光量,也有助於提高屈光系統的成像質量,瞳孔及視覺調節均受自主神經系統控制。
眼球的運動由六塊眼外肌來實現,這些肌肉的協調動作,保證了眼球在各個方向上隨意運動,使視線按需要改變。兩眼的眼外肌的活動必須協調,否則會造成視網膜雙像(復視)或斜視。
視網膜是一層包含上億個神經細胞的神經組織,按這些細胞的形態、位置的特徵可分成六類,即光感受器、水平細胞、雙極細胞、無長突細胞、神經節細胞,以及近年新發現的網間細胞。其中只有光感受器才是對光敏感的,光所觸發的初始生物物理化學過程即發生在光感受器中。
光感受器光感受器按其形狀可分為兩大類,即視桿細胞和視錐細胞。夜間活動的動物(如鼠)視網膜的光感受器以視桿細胞為主,而晝間活動的動物(如雞、松鼠等)則以視錐細胞為主。但大多數脊椎動物(包括人)則兩者兼而有之。在人的視網膜中,視錐細胞約有600~800萬個,視桿細胞總數達1億以上。它們似以鑲嵌的形式分佈在視網膜中;其分佈是不均勻的,在視網膜黃斑部位的中央凹區,幾乎只有視錐細胞。這一區域有很高的空間分辨能力(視銳度,也叫視力)。它還有良好的色覺,對於視覺最為重要。中央凹以外區域,兩種細胞兼有,離中央凹越遠視桿細胞越多,視錐細胞則越少。在視神經離開視網膜的部位(乳頭),由於沒有任何光感受器,便形成盲點。
視覺器官
視桿細胞的信號和視錐細胞的信號,在視網膜中的傳遞通路是相對獨立的,直到神經節細胞才匯合起來。在外網狀層,水平細胞在廣闊的範圍內從光感受器接收信號,並在突觸處與雙極細胞發生相互作用。此外,水平細胞還以向光感受器反饋的形式調製信號。在內網狀層雙極細胞的信號傳向神經節細胞,而無長突細胞則把鄰近的雙極細胞聯繫起來。視桿和視錐細胞信號的匯合也可能發生在無長突細胞。光感受器的信號主要通過改變化學性突觸釋放的遞質的量,向中間神經細胞傳遞。
視覺中樞的信息處理經過視網膜神經網路處理的信息,由神經節細胞的軸突——視神經纖維向中樞傳遞。
視覺器官
視覺信息在視覺中樞通路的各水平上經受進一步的處理。外側膝狀體只是視覺信息傳遞的中繼站。
眼的折光系統及其調節
當光線由空氣進入另一媒質構成的單球面折光體時,它進入物質的折射情況決定於該物質與空氣界面的曲率半徑R和該物質的折光指數n2;若空氣的折光指數為n1,則關係式為 n2R/(n2-n1)=F2 (1)
F2稱為後主焦距或第2焦距(空氣側的焦距為前主焦距或第一焦距),指由折射面到後主焦點的距離,可以表示這一折光的折光能力。表示折光體的折光能力還可用另一種方法,即把主焦距以m(米)作單位來表示,再取該數值的倒數,後者就稱為該折光體的焦度(diopter);如某一透鏡的主焦距為10cm,這相當於0.1m,則該透鏡的折光能力為10焦度(10D)。通常規定凸透鏡的焦度為正值,凹透鏡的焦度為負值。
主焦距是一個折光體最重要的光學參數,由此可算出位於任何位置的物體所形成的折射像的位置。以薄透鏡為例,如果物距α是已知的,像距b可由下式算出:
1/a+1/b=1/F2 (2)
由式(2)可以看出,當物距a趨於無限大時,1/a趨近於零,於是1/b接近於1/F2,亦即像距b差不多和F2相等;這就是說,當物體距一個凸透鏡無限遠時,它成像的位置將在後主焦點的位置。同樣不難看出,凡物距小於無限大的物體,它的像距b恆大於F2,即它們將成像在比主焦點更遠的地方。以上兩點結論,對於理解眼的折光成像能力十分重要。
視覺器官
眼的折光系統的光學特性
當用上述光學原理分析眼的折光特性時,首先遇到的一個困難是,眼球並非一個薄透鏡或單球面折光體,而是由一系列由率半徑和折光指數都不相同的折光體所組成的折光系統。顯然,人眼折光系統的後主焦距不能簡單地由式(1)算出,不過它的最主要的折射發生在角膜,而按幾何學原理進行較複雜的計算,還是可以追蹤出光線經眼內多個折光面行進的途徑,並得出由這些組合的透鏡組所決定的後主焦點的所在位置。計算結果表明,正常成人眼處於安靜而不進行調節的狀態時,它的折光系統的後主焦點的位置,正好是其視風膜所在的位置。這一解剖關係對於理解正常眼的折光成像能力十分重要。它說明,凡是位於眼前方6m以外直至無限遠處的物體,根據式(2)或由於由它們發出或反射出的光線在到達眼的折光系統時已近於平行,因而都可以在視網膜上形成基本清晰的像,這正如放置於照相機主焦點處的底片,可以拍出清晰的遠景一樣。當然,人眼不是無條件的看清任何遠處的特體,例如,人眼可以看清楚月亮(或其他更遠的星體)和它表面較大的陰影,但不能看清楚月球表面更小的物體或特徵。造成后一限制的原因是,如果來自某物體的光線過弱,或它們在空間處女內傳播時被散射或吸收,那麼它們到達視網膜時已減弱到不足以興奮感光細胞的程度,這樣就不可能被感知;另外,如果物體過小或它們離眼的距離過大,則它們在視網膜上形成的大小,將會小到視網膜分辨能力的限度以下,因而也不能感知。
眼的調節
如果安靜狀態的眼的折光能力正好把6m以外的物體成像在視網膜上,那麼來自較6m為近的物體的光線將是不同程度呈輻射狀的,它們在折射后的成像位置將在主焦點,亦即視網膜的位置之後;由於光線到達視網膜時尚未聚焦,因而物像是模糊的,由此也只能引起一個模糊的視覺形象。但正常眼在看近特時也十分清楚,這是由於眼在看近物時已進行了調節(accommodation),使進入眼內的光線經歷較強的折射,結果也能成像在視網膜上。人眼的調節亦即折光能力的改變,主要是靠晶狀體形狀的改變;這是一個神經反射性活動,其過程如下:當模糊的視覺形象出現在視區皮層時,由此引起的下行衝動經錐體束中的皮層-中腦束到達中腦的正中核,再到達發出動眼神經中副交感節前纖維的有關核團,最後再經睫狀神經節到達眼內睫狀肌,使其中環行肌收縮,引起連接於水晶體囊的懸韌帶放鬆;這樣就促使水晶體由於其自身的彈性而向前方和後方凸出(以前突較為明顯),使眼的總的折光能力較安靜時增大,使較輻射的光線提前聚焦,也能成像在視網膜上。表示調節前後晶狀體形狀的改變。很明顯,物體距眼球愈近,到達眼的光線輻散程度愈大,因而也需要晶狀體作更大程度的變凸。調節反射進行時,除晶狀體的變化外,同時還出現瞳孔的縮小和兩眼視軸向鼻中線的會聚,前者的意義在於減少進入眼內光線的量(物體移近時將有較強光線到達眼球)和減少折光系統的球面像差和色像差;兩眼會聚的意義在於看近物時物像仍可落在兩眼視網膜的相稱位置。
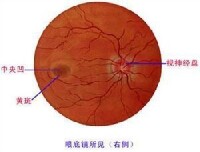
瞳孔指虹膜中間的開孔,是光線進入眼內的門戶;它在亮光處縮小,
視覺器官(右側眼球水平斷模式圖)在暗光處散大。虹膜由多單位平滑肌構成;在瞳孔周圍的是環形肌層,受動眼神經中的副交感神經纖維支配,收縮時使瞳孔縮小,故又稱瞳孔括約肌;虹膜的外周部分是輻散狀肌纖維,受由頸部上行的交感神經纖維支配,收縮時使瞳孔散大,故又稱瞳孔散大肌。瞳孔的大小可以控制進入眼內的光量。一般人瞳孔的直徑可變動於1.5-8.0mm之間。假定人由光亮處進入暗室時瞳孔直徑可增加5倍,那麼瞳孔的受光面積應增大25倍;可見瞳孔的變化,有保持在不同光照情況下進入眼內的光量較為恆定的作用。但暗室中較強陽光照射的光照強度實際減弱約100萬倍,因而單靠瞳孔大小的改變,遠不足以使進入眼內的光量保持恆定。事實上,人眼在不同的亮度情況下是靠視網膜中不同的感光細胞來接受光刺激的,在暗光處起作用的視桿細胞對光的敏感程度要比在亮光處起作用的視錐細胞大得多,因此在暗處看物,只需進入眼內光量適當增加即可。由此可見,通過改變瞳孔大小以調節進入眼內的光量還是有一定意義的。瞳孔大小隨光照強度而變化的反應,是一種神經反射,稱為瞳孔對光反射。引起此反射的感受器就是視網膜,傳入纖維在視神經中,但這部分纖維在進入中樞后不到達外側膝狀體,而在在中腦的頂蓋前區換神經元,然後到同側和對側的動眼神經核,傳出纖維主要是動眼神經中的副交感纖維,效應器也主要是瞳孔約肌。
瞳孔對光反應的特點是效應的雙側性,即如果光照的是一側眼睛時,除被照眼出現瞳孔縮小外,同時未受光照拐殊途同歸瞳孔也縮小,後者我為互感性對光反射。臨床上有時可見到瞳孔對光反應消失、瞳孔左右不等、互感性瞳孔反應消失等異常情況,常常是由於與這些反射有關的反射綿弧某一部分受損的結果,因而可以藉瞳孔反應的異常幫助進行神經病變的定位診斷。房水和眼內壓房水指充盈於眼的前、後房中的液體,其成分類似血漿,但蛋白質含量較血漿低得多,而HCO3-含量卻超過血漿;因而房水的總滲透壓也較血漿為高。房水的生成部位在睫狀體脈絡膜叢,生成後由後房經瞳孔進入前房,再由鞏膜和角膜結合處的前房角進入鞏膜靜脈竇,最後匯入靜脈系統。房水不斷生成,不斷回收入靜脈,使它在後房和前房之間流動不息。據測定,正常時房水的生成速度每分鐘約為2mm3;由於它的生成和回收之間保持著動態平衡,使得眼內政黨時的房水量保持恆定,又由於眼的前、後房容積也是相對恆定的,於是使其中靜水壓(即眼內壓)也保持相對的穩定。中國成年人眼內壓正常值為2.27-3.2kPa(17-24mmHg),平均2.67kPa(17-24mmHg)。眼內壓的相對穩定,對保持眼球特別是角膜的正常開頭和折光能力有重要的意義。當眼球被刺穿時,可能導致房水流失,眼內壓下降,引起眼球變形,角膜也不能保持正常的曲度。人眼的總折光能力與眼內折光體都有一定關係,但最主要的折射發生在空氣與角膜接觸的界面上。這約佔總折光能力的80%。因此,角膜的曲度和形狀的改變將顯著地影響眼的折光能力,嚴重地影響視力。房水也對它所接觸的無血管組織如角膜和晶狀體起著營養的作用。房水循環障礙時會造成眼內壓過高,臨床上稱為青光眼,可導致角膜、晶狀體以及虹膜等結構的代謝障礙,嚴重時造成角膜混濁、視力喪失。房水生成的機制目前尚不完全明了。
通常認為,除了在睫狀體脈絡膜叢處的毛細血管靠被動濾過(類似在一般毛細血管的動脈端生成組織液),使血漿中的水分和鹽類透出血管壁生成房水外,還有主動過程的參與,否則就難於解釋房水何以有較血漿中濃度高的HCO3-等鹽類離子。用組織化學的方法證明,睫狀體上皮細胞含有較多的碳酸酐酶,此酶的作用是使細胞代謝過程中產生的CO2和H2O,迅速生成,並解離出HCO3-,後者經膜上的主動轉運過程進入房水,造成它的房水中的高濃度,這個高濃度造成的負電位和高滲透壓還能進一步促使血漿中的Na 和水分子進入房水。臨床上可以使用碳酸酐酶抑製劑(如乙醯唑胺)減少房水生成,降低眼內壓,其作用機制與上述的房水生成機制有關。
來自外界物體的光線,通過眼內的折光系統在視網膜上形成物像,是視網膜內的感光細胞被刺激的前提條件。視網膜像還有一個物理範疇內的內像,用幾何光學的原理可以較容易地對它加以說明,和外界物體通過照相機的中的透鏡組在底片上形成的物像並無原則上的區別;但視覺系統最後在主觀意識上形成的“像”,則是屬於意識或心理範疇的主觀印象,它由來自視網膜的神經信息最終在大腦皮層等中樞結構內形成。作為感受器生理,重點是視網膜怎樣把物理像轉換成視神經纖維上的神經信號,以及在這些信號的序列和組合中怎樣包括了視網膜像、亦即外界物體所提供的信息內容。應該提出,視覺研究的進展雖然較快,但也只是初步的。
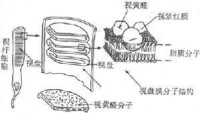
視網膜的厚度只有0.1-0.5mm,但結構十分複雜。它的主要部分在個體發生上來自前腦泡,故屬於神經性結構,其中細胞通過突觸相互聯繫。經典組織學將視網膜分為十層,但按主要的細胞層次簡化為四層業描述,從靠近脈絡膜的一側算起,視網膜最外層是色素細胞層;這一層的來源不屬神經組織,血液供應也來自脈絡膜一側,與視網膜其他層接受來自視網膜內表面的血液供應有所不同;臨床上見到的視網膜剝離,就發生在此層與其它層次之間。色素細胞層對視覺的引起並非無關重要,它含在黑色素顆粒和維生素A,對同它相鄰接的感光細胞起著營養和保護作用。保護作用是除了色素層可以遮繼來自鞏膜側的散射光線外,色素細胞在強光照射視網膜時可以伸出偽足樣突起,包被視桿細胞外段,使其相互隔離,少受其他來源的光刺激;只有在暗光條件下,視桿外段才被暴露;色素上皮的這種活動受膜上的多巴胺受體控制。此層內側為感光細胞層。在人類和大多數哺乳動作動物,感光細胞分視桿和視錐細胞兩種,它們都含有特殊的感光色素,是真正的光感受器細胞。視桿和視錐細胞在形態上都可分為四部分,由外向內依次稱為外段、內段、胞體和終足;其中外段是感光色素集中的部位,在感光換能中起重要作用。視桿和視錐細胞在形成上的區別,也主要在外段它們外形不同,所含感光色素也不同。視桿細胞外段呈長桿狀,視錐細胞外段呈圓錐狀。兩種感光細胞都通過終足和雙極細胞層內的雙極細胞發生突觸聯繫,雙極細胞一般再和節細胞層中的神經節細胞聯繫。視網膜中除了這種縱向的細胞間聯繫外,還存在橫向的聯繫,如在感光細胞層和雙極細胞層之間有水平細胞,大雙極細胞層和節細胞層之間有無長突細胞;這些細胞的突起在兩層細胞之間橫向伸展,可以在水平方向傳遞信息,使視網膜在不同區域之間有可能相互影響;這些無長突細胞還可直接向節細胞傳遞信號。近年來發現,在視網膜還存在一種網間細胞,它的細胞體位於雙極細胞層和節細胞層之間,但突起卻伸到感光細胞層和雙極細胞層。如果把感光細胞經過雙極細胞到神經節細胞的途徑,看作是視覺信息的初始階段。視網膜中除了有通常的化學性突觸外,還有大量電突觸存在。由此可見,視網膜也和神經組織一樣,各級細胞之間存在著複雜的聯繫,視覺信息最初在感光細胞層換能變成電信號后,將在視網膜複雜的神經元網路中經歷某種處理和改變,當視神經纖維的動作電位序列作為視網膜的最終輸出信號傳向中樞時,它們已經是經過初步加工和處理的信息了。
盲點由節細胞層發出的神經軸突,先在視網膜表面聚合成一整束,然後它透視網膜,在眼的后極出眼球,這就在視網膜表面形成視神經乳頭。在乳頭的範圍內,實際上沒有視網膜特有的細胞結構,因而落於該處的光線或視網膜像的組成部分,將不可能被感知,故稱為盲點。兩側視神經乳頭在視網膜內黃斑或中央凹中心的鼻側約3mm處。但正常時由於用兩眼看物,一側盲點可以被對側視覺補償,人們並不覺察自己的視野中有一處無視覺感受的區域。盲點的存在可用專門設計的方法來證明。
根據對視網膜結構和功能的研究,認為在人和大多數脊椎動物的視網膜中存在著兩種感光換能系統。一種由視桿細胞和與它們相聯繫的雙極細胞和神經節細胞等成分組成,它們對光的敏感度較高,能在昏暗的環境中感受光刺激而引起視覺,但視物無色覺而只能區別明暗;且視物時只能有較粗略的輪廓,精確性差,這稱為視桿系統或晚光覺系統;另一種由視錐細胞和與它們有關的傳遞細胞等成分組成,它們對光的敏感性較差,只有在類似白晝的強光條例下才能被刺激,但視物時可辨別顏色,且對物體表面的細節和輪廓境界都能看得很清楚,有高分辨能力,這稱為視錐系統或晝光覺系統(前述視敏度的測定實際是視錐系統視力的測定)。
證明這兩種相對獨立的感光-換能系統存在的主要依據是:
①人視網膜中視桿和視錐細胞在空間上的分佈是不均勻的,愈近視網膜周邊部,視桿細胞愈多而視錐細胞愈少;愈近視網膜中心部,視桿細胞愈少而視錐細胞愈多;在黃斑中心的中央凹處,感光細胞全部是視錐而無視桿細胞;與上述細胞分佈相對應,人眼視覺的特點正是中央凹在亮光處有最高的視敏度和色覺,在暗外則中央視力較差;相反地,視網膜周邊部則能感受弱光的刺激,但這時無色覺而清晰度較差。
②兩種感光細胞和雙極細胞以及節細胞形成信息傳遞通路時,逐級之間都有一定程度的會聚現象,但這種會聚在視錐系統程度較小,在中央凹處甚至可以看到一個視錐細胞只同一個雙極細胞聯繫,而這個雙極細胞也只同一個神經節細胞聯繫的情況,這種低程度會聚或無會聚現象的“單線聯繫”,顯然是視錐系統有較高的精細分辨能力的結構基礎;與此相對照,在視桿系統則普遍存在多個感光細胞同一個雙極細胞聯繫,而多個雙極細胞再同一個神經節細胞聯繫的會聚式排列;在視網膜周邊部,可看到多達250個視桿細胞經少數幾個雙極細胞會聚於一個神經節細胞的情況;在這種情況下,當然不能期待這樣的感覺系統有高的精細分辨能力。但這樣的聚合系統卻具有較強的總和多個弱刺激的能力。
③從動物各系統特點來看,某些只在白晝活動的動物如爬蟲類和雞等,視網膜全無視桿而只胡視錐細胞。而另一些只在夜間活動的動物如地松鼠和貓頭鷹等,視網膜中只胡視桿而不含視錐細胞。
④視桿細胞中只含有一種感光色素,即 視紫紅質(rhodopsin),而視錐細胞卻因所含感光色素的吸收光譜特性不同而分為三種,這是同視桿系統無色覺而視錐系統有色覺的事實相一致的。
有人就從視網膜中提取出了一定純度的感光色素即視紫紅質,它在暗處呈紅色;實驗中還可以證明,提取出來的這種感光色素對不同波長光線的吸收光譜,基本上和晚光覺對光譜不同部分的敏感性曲線相一致。這一事實十分重要,因為既然光線對某種感光色素的光化學作用的強度正好與這些光線所引起的視覺的強度相一致,那就是提示前者可能是後者的基礎。視紫紅質的光化學反應及其代謝 視紫紅質的分子量約為27-28kd,是一種與結合蛋白質,由一分子稱為視蛋白(opsin)的蛋白質和一分子稱為 視黃醛(retnal)的生色基團所組成。視蛋白的肽鏈序列已搞清,它的肽鏈中有7段穿越所在膜結構、主要由疏水性氨基酸組成的α-螺旋區段,同一般的細胞膜受體具有類似的結構。視黃醛由維生素A變來,後者是一種不飽和醇,在體內一種酶的作用下可氧化成視黃醛。提純的視紫紅質在溶液中對500nm波長的光線吸收能力最強,這與人眼在弱光條件下對光就業上藍綠光區域(相當於500nm波長附近)感覺最明亮(不是感到了藍綠色)的事實相一致,說明人在 暗視覺與視桿細胞中所含視紫紅質的光化學反應有直接的關係。視紫紅質對光譜不同部分的吸收曲線視覺中最明亮的區域和視紫紅制裁吸收能力最強的部分都在500nm的波長附近。視紫紅質在光照時迅速分解為 視蛋白和視黃醛,這是一個多階段的反應。分解的出現首先是由於視黃醛分子在光照時發生了分子構象的改變,即它在視紫紅質分子中本來呈11-順型(一種較為彎曲的構象),但在光照時變為全反型(一種較為直的分子構象)。視黃醛分子構象的這種改變,將導致視蛋白分子構象也發生改變,經過較複雜的信號傳遞系統的活動,誘發視桿細胞出現感受器電位。據計算,一個光量子被視紫紅質吸收,就足以使視黃醛分子結構發生改變,導致視紫紅質最後分解為視蛋白和視黃醛。視紫紅質分解的某些階段伴有能量的釋放,但這看來不是誘發感受器電位的直接原因。
在亮處分解的視紫紅質,在暗處又可重新合成,亦即它是一個可逆反應,其反應的平衡點決定於光照的強度。視紫紅質再合成的第一步,是全反型的視黃醛變為11-順型的視黃醛,很快再同視蛋白結合。此外,貯存在視網膜的色素細胞層中的維生素A也是全反型的,它們也可在耗能的情況下變成11-順型的,進入視桿細胞,然後再氧化成11-順型的視黃醛,參與視紫紅質的合成補充;但這個過程進行的速度較慢,不是促進視紫紅制裁再合成的即時因素。人在暗處視物時,實際是既有視紫紅質的分解,又有它的合成,這是人在暗光處能不斷視物的基礎;光線愈暗,全盛過程愈超過分解過程,視網膜中處於合成狀態的視紫紅質數量也愈高,這也使視網膜對弱光愈敏感;相反,人在亮光處時,視紫紅質的分解增強,合成過程甚弱,這就使視網膜中有較多的視紫紅質處於分解狀態,使視桿細胞幾乎失去了感受光刺激的能力;人的視覺在亮光處是靠另一種對光刺激較不敏感的感光系統即視錐來完成的,后一系統在弱光時不足以被刺激,而在強光系統下視桿細胞中的視紫紅質較多地處於分解狀態時,視錐系統就代之而成為強光刺激的感受系統。在視紫紅質和再合成的過程中,有一部分視黃醛被消耗,這最終要靠由食物進入血液循環(相當部分貯存於肝)中的維生素A來補充。長期攝入 維生素A不足,將會影響人在暗光處的視力,引起夜盲症。
超威結構和感受器電位的產生
視桿細胞外段的超威結構和感受器電位的產生
感光細胞的外段是進行光-電轉換的關鍵部位。視桿細胞外段具有特殊的超威結構,在外段部分,膜內的細胞漿甚少,絕大部分為一些整齊的重疊成層的圓盤狀結構所佔據,這圓盤稱為視盤。每一個視盤是一個扁平的囊狀物,囊膜的結構和細胞膜類似,具有一般的脂質雙分子層結構,但其中鑲嵌著的蛋白質絕大部分是視紫紅質,亦即視桿細胞所含的視紫紅質實際上幾乎全部集中在視盤膜中。視盤的數目在不同動物的視桿細胞中相差很大,人的每個視桿細胞外段中它們的數目近千;每一個視盤所含的視紫紅質分子約有100萬個。這樣的結構顯然有利於使進入視網膜的光量子有更大的機會在外段中碰到視紫紅質分子。
有人用細胞內微電極技術,研究了視桿細胞外段內外的電位差在光照前後的變化,結果發現在視網膜未經照射時,視桿細胞的靜息電位比一般細胞小得多。經分析表明,這是由於外段膜在無光照時,就有相當數量的Na 通道處於開放狀態並有持續的Na 內流所造成,而內段膜有Na 泵的連續活動將Na 移出膜外,這樣就維持了膜內外的Na 平衡。當視網膜受到光照時,可看到外段膜兩側電位短暫地向超極化的方向變化,由此可見,外段膜同一般的細胞膜不一致,它是在暗處或無光照時處於去極化狀態,而在受到光刺激時,跨膜電痊反而向超極化方向變化,因此視桿細胞的感受器電位(視錐細胞也一樣),表現為一種超極化型的慢電位,這在所有被研究過的發生器或感受器電位中是特殊的,它們一般都表現為 膜的暫時去極化。
光子的吸收引起外段膜出現超極化電反應的機制已基本搞清,這就是光量子被作為受體的視紫紅質吸收后引起視蛋白分子的變構,又激海參了視盤膜中一種稱為傳遞蛋白(transducin)Ct的中介物,後者在結構上屬於G-蛋白家庭的一員,它激活的結果是進而激活附近的磷酸二酯酶,於是使外段部分胞漿中的cGMP大量分解,而胞漿中cGMP的分解,就使未受光刺激時結合於外段膜的cGMP由也膜解離而被分解,而cGMP在膜上的存在正是這膜中存在的化學門控式Na 通道開放的條件,膜上cGMP減少,Na 通道開放減少,於是光照的結果出現了我們記錄到的超極化型感受器電位。據估計,一個視紫紅質被激活時,可使約500個傳遞蛋白被激活;雖然傳遞蛋白激活磷酸二酯酶是1對1的,但一個激活了的磷酸二酯酶在一秒鐘內大約可使4千多個cGMP分子降解。由於酶系統的這種生物放大作用,就可以說明1個光量子的作用何以能在外段膜上引起大量化學門控式Na 通道的關閉,引起一個足以為人的視覺系統所感知的超極化型電變化。
視桿細胞外段和整個視桿細胞都沒有產生動作電位的能力,由光刺激在外段膜上引起的感受器電位只能以電緊張性的擴布到達它的終足部分,影響終點(相當於軸突末稍)外的遞質釋放。
視錐系統外段也具有與視桿細胞類似的盤狀結構,並含有特殊的感光色素,但 分子數目較少。已知,大多數脊椎動物具有三種不同的視錐色素,各存在於不同的視錐細胞中。三種視錐色素都含有同樣的11-順型視黃醛,只是視蛋白的分子結構稍有不同。看來是視蛋白分子結構中的微小差異,決定了同它結合在一起的視黃醛分子對何種波長的光線最為敏感,因而才有視桿細胞中的視紫紅質和三種不同的視錐色素的區別。光線作用於視錐細胞外段時,在它們的外段膜兩側也發生現視桿細胞類似的超級化型感受器電位,作為光-電轉換的第一步。認為視錐細胞外段的換能機制,也與視桿細胞類似。
視錐細胞功能的重要特點,是它有辨別顏色的能力。顏色視覺是一種複雜的物理-心理現象,顏色的不同,主要是不同波長的光線作用於視網膜后在人腦引起的主觀印象。人眼一般可在光譜上區分出紅、橙、黃、綠、青、藍、紫等七種顏色,每種顏色都與一定波長的光線相對應;但仔細的檢查可以發現,單是人眼在光譜可區分的色澤實際不下150種,說明在可見光譜的範圍內波長長度只要有3-5nm的增減,就可被視覺系統分辨為不同的顏色。很明顯,設想在視網膜中存在上百種對不同波長的光線起反應的視錐細胞或感光色素,是不大可能的。但物理學上從牛頓的時代或更早就知道,一種顏色不僅可能由某一固定波長的光線所引起,而且可以由兩種或更多種其他波長光線的混合作用而引起。例如,把光譜上的七色光在所謂牛頓色盤上旋轉,可以在人眼引起白色的感覺;用紅、綠、藍三種色光(不是這三種顏色的顏料)作適當混合,可以引起光譜上所有任何顏色的感覺。這后一現象特別重要;這種所謂三原色混合原理不僅早已廣泛地應用於彩色照像、彩色電視等方面,而且被用於說明顏色視覺的產生原理本身。早在上世紀初,Young(1809)和Helmholtz(1824)就提出了視覺的三原色學說,設想在視網膜中存在著分別對紅、紅、藍的光線特別敏感的三種視錐細胞或相應的三種感光色素,並且設想當光譜上波和介於這三者之間的光線作用於視網膜時,這些光線可對敏感波長與之相近兩種視錐細胞或感光色素起不同程度的刺激作用,於是在中樞引起介於此二原色之間的其他顏色的感覺。視覺三原色學說用較簡單的生物感受結構的假設說明了複雜的色覺現象,一般為多數人所接受;但在實驗中試圖尋找出遊同種類的視錐細胞或感光色素長時間未獲成功。用光學顯微鏡和電子顯微鏡不能發現視錐細胞之間在結構上有什麼不同,同時也未能用一般的化學方法分離郵不同的視錐感光色素。
由於實驗技術的進步,關於視網膜中有三種對不同波長光線特別敏感的視錐細胞的假說,已經被許多出色的實驗所證實,例如,有人用不超過單個視錐直徑的細小單色光束,逐個檢查並繪製在體(最初實驗是在金公和蠑螈等動物進行,以後是人)視錐細胞的光譜吸收曲線,發現所有繪製出來的曲線不外三種類型,分別代表了三類光譜吸收特性不同的視錐細胞,一類的吸收峰值在420nm外,一類在531nm外,一類在558nm外,差不多正好相當於藍、綠、紅三色光的波長,和上述視覺三原色學說的假設相符。用微電極記錄單個視錐細胞感受器電位的方法,也得到了類似的結果,即不同單分光引起的超極化型感受器電位的大小,在不同視錐細胞是不一樣的,峰值出現的情況符合於三原色學說。
三原色學說和它的實驗依據,大體上可以說明臨床上遇到的所謂色盲和色弱的可能發病機制。紅色盲也稱第一色盲,被認為是由於缺乏對較長波長光線敏感的視錐細胞所致;此外還有綠色盲,也稱第二色盲,藍色盲也稱第三原色盲,都可能是由於缺乏相應的特殊視錐細胞所致。紅色盲和綠色盲較為多見,在臨床上都不加以區別地稱為紅綠色盲;藍色盲則極少見。色盲患者的顏色不僅不能識別綠色,也不能區分紅也綠之間、綠與藍之間的顏色等。有些色覺異常的人,只是對某種顏色的識別能力差一些,亦即他們不是由於缺乏某種視錐細胞,而只是後者的反應能力較正常人為弱的結果,這種情況有別於真正的色盲,稱為色弱。色盲除了極少數可以由於視網膜後天病變引起外,絕大多數是由遺傳因素決定的。
三原色學說雖然比較圓滿地說明許多色覺現象和 色盲產生的原因,並已在光感受細胞的一級得到了實驗證實,但並不能解釋所有的顏色視覺現象,如顏色對比現象就是一個例子。試將藍色的小紙塊放在黃色或其它顏色的背景上,會覺得放在黃色背景上那個藍紙塊特別藍,同時覺得背景也比未放藍紙塊時更黃(在中國北方的黃土高原,當春天的風造成黃塵蔽日的情況時,會覺得平常的日晃燈管的 光線變得較藍了)。這種現象稱為顏色對比,而黃和藍則稱為對比色或互補色。顏色對比現象只出現對比色之間,而不是任意的兩種顏色之間。互為對比色的顏色對尚有:紅一綠以及黑和白。根據顏色對比等不容易用三原色學說圓滿視覺現象,幾乎是在三原色學說提出的同時就出現了另一種色覺學說,稱為對比色學說。該學說提出在視網膜中存在著三種物質,各對一組對比色的刺激起性質相反的反應。如前所述,近年來在視錐細胞一級進行的研究有利於三原色學說而不利於對比色學說,但後來在視網膜其它層細胞進行的一些實驗卻又符合對比色學說的推測。如在金魚水平細胞進行的微電極研究說明,此類細胞和視桿、視錐細胞不同,既能出現超極化的跨膜 電位改變,也能出現去極化型的電位改變,而且在用多種不同色光刺激時發現,有些水平細胞在黃光刺激時出現最大的去極化反應,在藍光刺激時出出現最大的超極化型反應;另一些水平細胞則在紅和綠色刺激時有類似的不同反應。這些現象是同對比色學說一致的。看來可能的是,各以部分色覺現象為出發點的兩種色覺學說都是部分正確的,在視錐細胞一級,不同色光以引起三種不同視錐細胞產生不同大小的超極化型電變化進行編碼;但到了水平細胞一級或其它級細胞(包括某些中樞神經元),信息又進行重新編碼,不同顏色雙可以用同細胞對互為對比色的顏色出現相反形式的電反應來編碼。以上事實說明,顏色視覺的引起是一個十分複雜的過程,它需要有從視網膜視錐細胞到皮層神經元的多級神經成分的參與才能完成。
由視桿和視錐細胞產生的電信號,在視網膜內要經過複雜的細胞網路的傳遞,最後才能由神經節細胞發生的神經纖維以動作電位的形式傳向中樞。由於視網膜內各種細胞之間的排列和聯繫非常複雜,與細胞間信息傳遞有關的化學物質種類繁多(除一般神經系統中常見的遞質外,連同視網膜中已發現的各神經肽類物質,總數已達30餘種),因而視覺信息在從感光細胞向節細胞傳遞時,必然要經歷種種改變;這實際就是視網膜本身對視覺信息的初步處理,它是在視網膜特定的細胞構築和化學構築的網路中按照某些規律進行的,但對這些規律的了解還是很膚淺的。現在所能初步肯定的是,雙極細胞、水平細胞和多數無長突細胞也同兩種感光細胞一樣,沒有產生動作電位的能力(但部分無長突細胞可產生動作電位);但這三種細胞同感光細胞不同的是,它們在前一級細胞的影響之下,既能產生超級化型慢電位,也能產生去極化型慢電位(相當於一般神經元突觸后膜處的IPSp和EPS)。所有這些慢電位,只能作電緊張性的擴布,影響突觸前膜遞質釋放量的改變,從而引起下一級細胞產生慢電位變化(也包括電突觸性相互影響);只有當這樣的慢電位傳遞到神經節細胞體時,由於後者有產生動作電位的能力,當兩種形式的慢電位總和的結果,使節細胞的靜息膜電位能夠去極經到閾電位水平時,才會產生“全或無”式的動作電位,作為視網膜的最後輸出信號傳向中樞。只有神經節細胞能產生動作電位雖然關於視網膜中信息處理的細節還不很清楚,但不妨先把視網膜作為控制論中的“黑箱”來看待,只對它的輸入(相當於作用於視網膜的光刺激)和它的輸入(相當於由視神經傳出的動作電位序列)進行對照分析,這樣也可以初步理解視網膜信息處理和編碼的最終結果。首先要記信一個事實是,視神經中纖維的總數(亦即節細胞的總數),只有全部感光細胞的1%。這一簡單事實就足以說明,視神經不可能通過其纖維“點對點”地傳遞視網膜中各感光細胞被光照的情況(中央凹處少數視錐細胞例外);因而大多數視神經纖維所傳遞的信號,只能是決定於多個感光細胞並因而含較多的信息量。
用小的光點刺激貓視網膜並同時記錄單條視神經纖維動作電位的方法表明,發出視神經纖維的節細胞大致可分為三類,分別稱為X-、Y-和W-細胞。X-和Y-細胞的特點是它們都具有大致呈同心圓式的“中心-周邊感受野”;一個節細胞的感受野在這裡指視網膜上某一特定的區域,當後者受到刺激時可能使該節細胞發生反應;但這兩種細胞的感受野都由兩部分構成;當光線作用於感受野的中心部分時出現節細胞放電增加,而當光線作用於環繞該中心的一定範圍內視網膜部分時,反而使該節細胞放電減少,這是一種類型的中心-周邊感受野,這就是當感受野的中心部分接受光刺激時引起節細胞放電數減少,而刺激其周邊部分時,反而引起該細胞放電增多,這種節細胞可稱為中心撤光反應細胞。
X-和Y-細胞都可以有上述兩種類型的感受野。它們之間的區別是,X-細胞的感受野較小,對刺激的反應是持續的;Y-細胞的感受野較大,而反應是時相性的,與刺激呈非線性關係。在猴子的實驗中還發現,不同的X-細胞對不同波長的光線反應不同,但對光的強度變化不敏感;而Y-細胞則對光照的強度變化敏感,卻對光線的波長變化不敏感。至於W-細胞,它們有較前兩者大得多的感受野,或是在光刺激時有放電,或是在撤光時有放電,但對刺激的反應比較遲緩,一般沒有性質相反的周邊視野區域。從形態學上看,X-和Y-細胞主要由雙極細胞接受輸入,而W細胞則主要接受無長突細胞的傳入。
每一條視神經纖維上的神經衝動並不簡單地表示某一部分視網膜受到光照或無光照,以上述X-為例,它的最強烈放電是出現在其感受野中心部分受到光照、而其外周部分全無光照(或相反的情況)的時候;如果中心和外周部同時受到光照,此節細胞的放電反而無大變化或只少有增加。另外,有事實說明,視網膜像經視網膜處理后,已被分解為不同“像素”,如有節細胞貢中樞傳輸圖像組成波長的住處有的傳輸它的不同亮度。這種把感受信息分解為其組成的“要素”,再在感受通路中進行“平行”傳輸和處理,在腦的各種感覺功能的研究中經常遇到,但是人的視覺也和另的感覺一樣,最複雜的信息處理和加工發生在中樞、特別是它的高級部分。
暗適應和明適應
人從亮處進入暗室時,最初看不清楚任何東西,經過一定時間,視覺敏感度才逐漸增主,恢復了在暗處的視力,這稱為暗適應。相反,從暗處初來到亮光處,最初感到一片耀眼的光亮,不能看清物體,只有稍待片刻才能恢復視覺,這稱為明適應。
暗適應是人眼對光的敏感度在暗光處逐漸提高的過程。在進入暗室后的不同時間,連續測定人的視覺閾值,亦即測定人眼剛能感知的光刺激強度,可以看到此閾逐漸變小、亦即視覺的敏感度在暗處逐漸提高的過程。一般是在進入暗室后的最初約7分鐘內,有一個閾值的明顯下降期,以後又出現閾值的量明顯下降;於進入暗室后的大約25-30分鐘時,閾值下降到最低點,並穩定於這一狀態。暗適應的產生機制與視網膜中感光色素在暗處時再合成增加,因而增加了視網膜中處於未分解狀態的色素的量有關。據分析,暗適應的第一階段主要與視錐細胞色素的合成量增加相一致;第二階段亦即暗適應的主要構成部分,則與視桿細胞中視紫紅質的合成增強有關。
明適應出現較快,約需一分鐘即可完成。耀眼的光感主要是由於在暗處蓄積起來的合成狀態的視紫紅質在進入亮處時先迅速分解,因為它對光的敏感性較視錐細胞中的感光色素為高;只有在較多的視桿細胞色素迅速分解之後,對光較不敏感的視錐細胞色素才能在亮光環境中感光。
視野
單眼固定地注視前方一點不動,這時該眼所能看到的範圍稱為視野。視野的最大界限應以它和視軸(單眼注視外界某一點時,此點的像正好在視網膜黃斑中央凹處,連接這兩點的假想線即視軸)所成夾角的大小來表示。在同一光照條件下,用不同顏色的目標物測得的視野大小不一樣,白色視野最大,其次為黃藍色,再次為紅色,而以綠色視野為最小。設想視野的大小除與各類感光細胞在視網膜中的分佈範圍有關外,在細節上還缺乏圓滿的解釋。另外,由於面部結構阻擋視線,也影響視野的形狀,如一般人顳側視野較大,鼻側視野較小等。臨床醫生檢查視野,使用特製的視野儀,並用不同顏色的視標進行檢查,目的在於了解視網膜的普遍感光能力,有時可藉以發現較大範圍的視網膜病變。某些視網膜、視神經或視覺傳導路的病變,有特殊形式的視野缺損,在診斷有意義。
視網膜電圖
將一個引導電極和角膜接觸,另一個電極置於額部作為參考電極,當給視網膜以廣泛光刺激時,可以在靈敏的電測量儀器上記錄到一系列電變化,這稱為視網膜電圖(electroretinogram)。視網膜電圖在性質上不同於同微電極在單一視網膜細胞成分記錄到的電現象;視網膜電圖是整個視網膜中各種成分在受到大範圍光照時的多種電反應的綜合反映。它通常由命名為a、b和c的三個波組成。據實驗分析,a波主要來源於感光細胞的感受器電位;b波幅度較大,主要與雙極細胞等細胞的活動有關;c波平緩而持續時間長,可能與色素細胞層的活動有關。有時在光照撤除時還可在緩慢持續的c波上再出現一個波動,稱為d波,產生原因尚不明了。視網膜電圖雖易於測量和描記,但反映視網膜功能狀態或病變的特異性不大,目前只發現少數疾病時有特殊的視網膜電圖改變,故在臨床上作用的意義不很大。
雙眼視覺和立體視覺
人和高等哺乳動物的雙眼都在面部前方,視物時兩眼視野差不多部分的像又各循自己特有的神經通路傳向中樞,但正常人主觀察感覺上只產生一個“物”的感覺。兩眼視物而只產生一個視覺形象的前提條件是:由物質同一部分的的光線,應成像在兩側視網膜的相稱點上。例如,兩眼的黃斑部就互為相稱點;當兩眼注視培牆上一個小黑點時,由於有眼外肌的調節,此點就都正好成像在兩側眼的黃斑上,於是在視覺中只“看到”一個點;此時如用手輕推一側眼球的外側,使此眼視軸稍作偏移,則這時此眼視網膜上的黑點像就要從黃斑部移開,落在與對側視網膜像非相稱的點上,於是會感到牆上有兩個黑點存在,這就是復視現象。顯然,在黃斑部以外,一眼的顳側視網膜和另一眼的鼻側視網膜互相對稱;而一眼的鼻側視網膜也與另正好的顳側視網膜互相對稱。
基本信息
- 中文名
- 視覺器官
- 外文名
- Визуальный орган
- 拼音
- shì jiào qì guān
- 定義
- 人和動物利用光的作用感知外界事物的感受器官
- 類型
- 醫學
- 別名
- eyespot
- 韓文名
- 시각 기관
- 拉丁文名
- visual organo
- 英文名
- Visual organ